


ABSTRACT
Cerebral ischemia is a major neurological condition that contributes significantly to global morbidity and mortality. It occurs due to reduced blood supply to the brain, leading to neuronal injury and death. The pathophysiology of cerebral ischemia is complex and involves oxidative stress, neuroinflammation, excitotoxicity, mitochondrial dysfunction, and apoptosis. Despite advances in medical management, effective neuroprotective therapies remain limited, creating a need to explore alternative approaches. Mucuna pruriens is a medicinal plant widely used in traditional medicine, known for its rich content of bioactive compounds, including levodopa, flavonoids, alkaloids, and phenolic antioxidants. These compounds are associated with antioxidant, anti-inflammatory, and neuroprotective properties. This review summarizes the available evidence from animal model studies on the role of
Keywords: Cerebral ischemia, Ischemic stroke, Mucuna, Neuroprotective agents.
INTRODUCTION
In 2021, 7.8 million people were affected by ischemic stroke globally, and 3.6 million people died during this period [1, 2]. Ischemic stroke comprises approximately 80% of all strokes and occurs because of reduced blood supply to the brain, causing damage to and death of brain tissue. There is no effective treatment for ischemic brain damage, possibly because of complex events that include excitotoxicity, loss of calcium homeostasis, oxidative stress, inflammation, lipid peroxidation, and apoptosis [3, 4], leading to neuronal death. Apoptosis is considered one of the critical factors responsible for postischemic cell death [5]. In global cerebral ischemia, the central part of the ischemic region does not receive any blood supply, but a small amount of blood can still reach this area through collateral circulation. The best example of this is middle cerebral artery occlusion-induced ischemia [6]. Reactive oxygen species (ROS) are generated after ischemia, leading to oxidative stress, neuronal death, and brain damage [7]. Newly formed free radicals extract an electron from another molecule, thus producing a chain reaction [8]. The synthesis of antioxidants balances the continuous production of oxidants. An imbalance between reactive oxygen metabolite production and antioxidant defenses results in oxidative stress, as shown in previous studies [9]. The accumulation of hydrogen peroxide impairs mitochondrial function [10], which persists for a longer duration after reperfusion and induces neuronal damage. There is a reduction in superoxide dismutase (SOD) and catalase (CAT) activity after ischemic injury. Apoptosis and necrosis are the two types of cellular death that occur after ischemia. In the core region, necrosis is more common, whereas neuronal death in the penumbral region is predominated by apoptosis [11]. Ischemia involves dysfunction of the hippocampal formation, neocortex, and Purkinje cells of the cerebellum. The hippocampus plays a major role in learning and memory. Compared with the CA3 region and dentate gyrus, the hippocampal CA1 region is more vulnerable to ischemic insult, and damage to this region is greater [12].
Fluoride, along with small amounts of aluminum, affects calcium influx and mobilization, neurotransmission, cell growth, differentiation, and cytoskeletal proteins [13]. Aluminum fluoride can affect learning and memory [14]. Various factors are involved, including synaptic plasticity because of the inhibition of long-term potentiation through the phosphoinositide 3-kinase–protein kinase B–mammalian target of rapamycin and brain-derived neurotrophic factor–tropomyosin receptor kinase B pathways [15, 16]. Disruption of the balance between mitochondrial fission and fusion leads to mitochondrial dysfunction in ischemic stroke [17]. Together, activation of toll-like receptor 4, increased ROS, and neuroinflammation lead to oxidative stress [18]. Inhibition of the histone demethylase PHF8 and decreased brain-derived neurotrophic factor eventually lead to epigenetic modifications [19]. All these mechanisms of action affect long-term memory [20].
Cerebral ischemia is a major neurological condition that causes morbidity, disability, and mortality worldwide. This condition requires immediate attention and emergency treatment; however, despite the best available advanced investigations and management methods, there are issues such as a lack of early diagnosis, time limitations in administering treatment, and unclear pathophysiology and adverse effects, which include oxidative stress, neuroinflammation, and apoptosis. There is a need for alternative drugs that can exert neuroprotective effects.
There is no universally accepted benchmarking of phytochemical profiles and pharmacological activity in
The present review aims to provide a comprehensive and critical synthesis of the available preclinical evidence on the neuroprotective potential of
REVIEW METHODOLOGY
The methodology for this narrative review was designed to include a comprehensive and critical overview of the literature. The results of the comparative analysis of animal model studies of
Table 1. Comparative analysis of animal model studies of
| Authors | Animal | Mode of experimental model | Extract of | Biochemical markers studied | Histopathological observation | Behavioral analysis |
|---|---|---|---|---|---|---|
| Chandran et al. [21] | Wistar albino rat | Spinal cord injury model | Ethanolic | Malondialdehyde, superoxide dismutase (SOD), catalase (CAT) | Decrease in neuronal death, axon sprouting, and glial scarring | No significant improvement |
| Bhosle and Wadher [22] | Wistar albino rat | Chemical ischemia-induced by 3-nitropropionic acid | Ethanolic | SOD, CAT, glutathione peroxidase (GPx) | Preservation of neuronal integrity | Improved motor function |
| Nayak et al. [23] | Wistar albino rat | Bilateral common carotid artery occlusion | Methanolic | SOD, CAT, GPx | Neuro-restoration, increased number of viable neurons | Improved locomotion, coordination of movements, and spatial learning |
| Dogra et al. [24] | Zebrafish | Rotenone-induced neurodegeneration | Chemically standardized | CAT, glutathione-S-transferase, acetylcholinesterase, butyrylcholinesterase | Decrease in inflammation and demyelination, reduction in focal necrosis in parts of the brain | Regaining of interest and cognition |
| Yadav et al. [25] | Swiss albino mice | 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced Parkinson’s | Ethanolic | Dopamine, 3,4-dihydroxyphenylacetic acid, homovanillic acid | Increase in the number of glial fibrillary acidic protein-positive neurons and astrocytes, recovery of tyrosine hydroxylase-positive neurons | Improved neurobehavioral performance |
| Manyam et al. [26] | Sprague–Dawley rat | 6-Hydroxydopamine-lesioned Parkinson’s model | Powdered seed, nonextracted form | Mitochondrial complex-I | Restoration of levodopa, dopamine, norepinephrine, and serotonin in the substantia nigra | Decreased symptoms of Parkinsonism |
Literature search strategy
The search was performed through electronic databases such as PubMed, Scopus, Web of Science, and Google Scholar. Studies published between 2000 and 2026 were given priority. The Medical Subject Headings browser and keyword search were used. The key search terms included “cerebral ischemia,” “
Article selection criteria
Initially, articles were selected based on their titles and abstracts. This was subsequently followed by downloading the full texts to read the manuscripts completely.
Inclusion criteria
In animal model studies of cerebral ischemia, studies evaluating any extract of
Exclusion criteria
Editorials, nonischemic models, non-extraction studies of
Screening procedure
Three authors reviewed the articles included in the literature search.
Complexity of ischemic brain injury
Ischemic brain injury involves complex biochemical, molecular, and cellular mechanisms. The cascade of events includes necrosis, apoptosis, the autoimmune response, and neuroplasticity [27]. Age-based variation has been reported to affect neuroprotection and ischemic outcomes [28]. Increased levels of neurotransmitters, including glutamate, contribute to excitation of receptors and subsequent neuronal injury. Disturbance in calcium ion homeostasis aggravates cell injury [29]. Astrocytic and microglial activation, along with infiltration of neutrophils and lymphocytes, leads to amplification of the inflammatory response [30]. The pathophysiology of cerebral ischemia is represented in Figure 1 [3, 5, 7, 8, 11, 23].
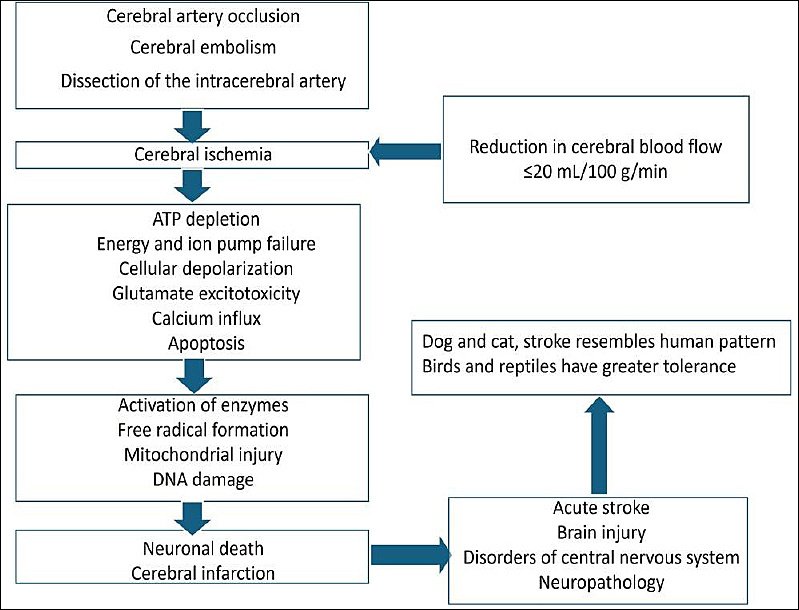
Figure 1. Schematic diagram showing the pathophysiology of cerebral ischemia [3, 5, 7, 8, 11, 23].
Research using animal models of ischemia suggests that reactive gliosis results from a response to altered perfusion. Changes in blood flow lead to glial activation and subsequent gliosis [31]. However, when chronic reactive gliosis occurs, neuronal recovery and regeneration are inhibited. Chronic astrogliosis is associated with the formation of glial scarring, which creates physical and molecular barriers that impede axon regeneration and functional recovery [32]. Microglial activation also triggers astrogliosis through the release of cytokines, which can exacerbate astrocytes’ reactive state, furthering neuronal injury.
Ischemic cascade
Immediately after the reduction in blood flow, oxygen and glucose levels decrease, further reducing adenosine triphosphate and energy-dependent processes [33]. Chronic cerebral ischemia occurs because of long-term reduction in cerebral blood flow caused by conditions such as hypertension and atherosclerosis, resulting in impaired cognition and dementia. Ionic imbalance occurs with the influx of sodium and calcium into cells and the involvement of the mitochondrial permeability transition pore. Nitric oxide and peroxynitrite also play important roles in the etiopathogenesis of ischemia–reperfusion injury.
Reperfusion injury in cerebral ischemia
Restoration of the blood supply can paradoxically aggravate injury through oxidative stress, inflammation, and disruption of the blood–brain barrier [30]. Leukocytic infiltration and activation of platelets and the complement system play instrumental roles [34]. Ischemia followed by reperfusion can affect mitochondria, leading to dysfunction. Injury to the mitochondrial complex and oxidative modifications are crucial aspects of the pathogenesis of ischemic neuronal injury [35]. The NLRP3 inflammasome and the Nrf2–Keap1 pathway are important during the management of reperfusion injury.
Autophagy and the neuroinflammatory cascade
Autophagy impairment eventually leads to the accumulation of damaged organelles and protein misfolding and aggregation [36]. The proinflammatory (M1) and anti-inflammatory (M2) phenotypes of microglial polarization play crucial roles in neuroinflammation [30]. Release of chemokines and inflammatory cytokines further contributes to mediation of neuronal injury and disruption of the blood–brain barrier. The LC3-II, Beclin-1, and p62 pathways, crosstalk between autophagy and apoptosis, M1/M2 ratios, and specific microglial marker TLR4/NF-κB pathways are also important. Rapamycin and 5-aminoimidazole-4-carboxamide ribonucleotide are autophagy activators that can increase neuronal viability, reduce lactate dehydrogenase leakage, and alleviate apoptosis in oxygen–glucose deprivation/reoxygenation models by activating the adenosine monophosphate-activated protein kinase/DDiT4/mammalian target of rapamycin axis [37].
Nuclear factor erythroid 2-related factor 2 (Nrf2)
Nrf2 is a regulator of the antioxidant response at the cellular level and activates the expression of proteins and enzymes, which help neutralize ROS-induced ischemia–reperfusion injury. Nrf2 helps maintain mitochondrial function, biogenesis, and integrity, which are essential for neuronal survival during ischemia. Its activation can inhibit iron-dependent lipid peroxidation in the context of cerebral ischemia [38, 39].
Cerebral ischemia in an animal model
In the global ischemic model, there is complete disruption of blood flow to the brain, leading to necrosis of the cerebral region. The methods used to achieve global cerebral ischemia include increasing intracranial pressure, occlusion of major arteries, and cervical compression [40]. However, a straightforward method for inducing ischemia is decapitation, which is performed after approval from the institutional animal ethics committee. This method was used long ago in small animals to elucidate the biochemical mechanism and global ischemic pathways involved. In the global cerebral ischemic model, any artery supplying the cerebral region can be occluded. The best example is middle cerebral artery occlusion in small-animal or large-animal model studies [41]. The middle cerebral artery occlusion model is widely used because it simulates thromboembolic stroke. Ischemic models can be generated in both small and large animals. Although many drawbacks exist in using animal models to study ischemia, these models are still in use.
There may be species-specific variations in neuroanatomy and neurophysiology, including responses to ischemia. The size and structure of the brain, vasculature, neuroprotection, and metabolic responses can vary among rat models, larger animals, and humans. Induction of cerebral ischemia by occluding the middle cerebral artery can produce variable results because of differences in surgical techniques, methods of anesthesia, and postoperative care. While animal model studies can yield good results, variable outcomes can cause difficulty in interpretation of the findings. Factors such as the genetic background of the animals can affect the severity of ischemic events and the recovery phase. Therefore, the findings of animal model research cannot be generalized. However, physiological control can be achieved in these models. With the help of these models, injury, its mechanism, and its neuroprotective effects can be better understood. These phases can be challenging to study in humans during ischemia because of the varied causes, manifestations, and ischemic sites. However, studying the mechanism of injury as well as neuroprotection is highly beneficial.
Nayak et al. [23] induced cerebral ischemia through bilateral carotid artery occlusion and thereby created oxidative stress. Ischemia results in a decline in memory and learning abilities; later, locomotor activity was assessed using an actophotometer, and motor coordination was assessed using the rotarod test. This was supported by assessment of cerebral infarction on the basis of histopathological findings. This was further supported by biochemical evidence of elevated lipid peroxidase (LPO) levels and depletion of SOD, CAT, reduced glutathione (GSH), and total thiols in the ischemic group. They also chemically induced ischemia by inducing oxidative stress through the administration of aluminum fluoride in drinking water for 7 days. There were decreased levels of SOD, CAT, and glutathione peroxidase (GPx), and elevated levels of oxidative stress markers, including malondialdehyde (MDA) and ROS.
Increased intracranial pressure (ICP) and cerebral ischemia
The standard ICP threshold is more than 20 mmHg, and it can be measured with modern ICP monitoring devices, which may be invasive. Increased ICP can result from space-occupying lesions, cerebral edema, and hematomas. This reduces cerebral perfusion pressure and cerebral blood flow, leading to global ischemia and hypoxia. It causes mechanical distortion and compression of the brain, exacerbating ischemia [42]. A reduction in cerebral blood flow can lead to metabolic dysfunction and secondary brain injury. Research has suggested a significant decrease in cortical blood flow and velocity because of increased ICP [43]. In animal models of traumatic brain injury, increased ICP leads to increased neuronal injury and behavioral changes, even without global ischemia. This finding indicates that even subischemic increases in ICP can cause significant deficits [44].
Experimental induction of increased ICP in animal models
Rodents, canines, pigs, and nonhuman primates have been used as animal models to induce increased ICP. Conditions such as an acute subdural hematoma cause increased ICP and exacerbation because of the mass effect of the hematoma. However, experimental animal studies often involve the infusion of artificial lumbar cerebrospinal fluid to increase ICP [43, 45]. In a Japanese study, autologous arterial blood was infused into the brainstem and internal capsule to increase ICP in white rabbits [46]. In another study, a Fogarty balloon catheter was inserted and inflated to simulate a space-occupying lesion and increased ICP [47].
Injection of autologous blood into the midbrain, internal capsule, and related sites can cause hemorrhage and increased ICP. Injection of saline into the lateral ventricle or brain can also cause increased ICP. In pigs, this method involves monitoring the optic nerve sheath diameter by ultrasound, which is an indirect indicator of increased ICP [48]. Direct measurement of ICP using transducers placed in the brain parenchyma or lateral ventricle is accurate but technically difficult [48].
Institutional animal ethics committee approval is mandatory to induce increased ICP in animals. It is also suggested that the induction and measurement of increased ICP can be complex due to the need for specialized equipment and expertise, potentially leading to variable results.
Induction of cerebral ischemia in animals by occlusion of arteries
Cerebral ischemia can be experimentally induced through various arterial occlusions. The middle cerebral artery can be occluded by electrocoagulation through the application of an electric current to coagulate the artery. Devices such as filaments can be used and adjusted to create either permanent or transient occlusions of the middle cerebral artery. Application of the endothelin-1 peptide also induces vasospasm, leading to ischemia [49]. The common carotid artery can also be occluded to induce ischemia. Occlusion of the artery leads to decreased nutrition, reduced energy production, an acidic environment, and resultant cell death [50]. This method requires strict aseptic precautions and physiological monitoring under anesthesia. The limitation of this method is the presence of collateral circulation, which can significantly influence the outcome.
Middle cerebral artery occlusion models are commonly used in stroke research to simulate ischemic conditions in the brain. These models can be categorized into permanent and transient occlusion models. Permanent occlusion is usually induced by electrocoagulation, which permanently blocks the artery without reperfusion. Transient occlusion involves temporary occlusion of the artery with an intraluminal suture, followed by reperfusion after a period ranging from 30 min to 2 h. The Longa technique involves generating a focal ischemia model by occluding the right middle cerebral artery. The animals were excluded if a subarachnoid hemorrhage occurred during middle cerebral artery occlusion [51].
Treatment of cerebral ischemia in an animal model
Given that ROS play a significant role in cell death following ischemic injury, several studies have been conducted to assess their neuroprotective effects. Modulation of hypoxia-inducible factor alpha has also been shown to increase endothelial nitric oxide synthetase and enhance cerebral blood supply in animal models of arterial occlusion [52]. Although this concept has shown positive results in animal models, in human trials these findings have not been as robust. This concept is successful in cardiovascular surgery but not in neurological disorders. The systematic use of imaging in ischemia and its management will allow us to understand the exact phenomenon occurring in the brain [53]. Therefore, there is a need to use combined approaches in ischemic conditions, including neuroprotective and vasoprotective agents, together with revascularization treatments.
The search for a natural neuroprotective substance with minimal side effects has attracted increasing attention. Although many plants have been examined to date, only a few have been studied thoroughly. Various compounds with antioxidant properties have been studied and tested to assess their endogenous antioxidant effects. Free radical scavengers and antioxidants are commonly used to mitigate the harmful effects of oxygen-free radicals during ischemia. There are potential clinical benefits, including reduced histopathological, biochemical, and oxidative metabolic damage, as well as improved neurological outcomes. A comparison of various drugs used to manage cerebral ischemia in animal models is presented in Table 2 [54–61].
Table 2. Comparison of various drugs used in the management of cerebral ischemia in animal models.
| Drug | Class | Mechanism of action | Experimental model | Neuroprotective effects | Dose | Translational status |
|---|---|---|---|---|---|---|
| Dizocilpine | N-methyl-D-aspartate receptor antagonist | Noncompetitive blockade of N-methyl-D-aspartate receptors → decreased Ca²⁺ influx → decreased excitotoxic neuronal death | Middle cerebral artery occlusion-induced global ischemia in rat, mouse, and cat | Decreased infarct volume, decreased neuronal death, improved motor and cognitive outcomes | Pre- or early postischemia (0–2 h); 0.1–5 mg/kg | Strong preclinical efficacy but failed clinical translation because of psychotomimetic and neurotoxic side effects [54] |
| Edaravone | Free radical scavenger | Scavenges ROS, activates Nrf2/antioxidant response element pathway, inhibits lipid peroxidation, ferroptosis, and apoptosis | Middle cerebral artery occlusion-induced global ischemia in rat | Decreased infarct size, decreased oxidative stress, decreased inflammation, increased brain-derived neurotrophic factor, improved neurological scores | Postischemia (≤6 h); 3–30 mg/kg | Clinically approved in Japan; strong alignment between animal and clinical data [55] |
| Edaravone–dexborneol | Antioxidant + anti-inflammatory combination | Inhibits oxidative stress, NF-κB/NLRP3, ferroptosis, and pyroptosis; enhances blood–brain barrier integrity | Middle cerebral artery occlusion-induced global ischemia in rat | Decreased infarct volume, increased cerebral blood flow, decreased neuroinflammation, improved cognition | Immediate to delayed postischemia; 0.375–15 mg/kg | Improved efficacy over edaravone alone; high translational potential [56] |
| Nimodipine | L-type Ca²⁺ channel blocker | Blocks voltage-gated Ca²⁺ channels, causes vasodilation, and decreases ischemic acidosis | Middle cerebral artery-induced hypertensive rat models | Decreased infarct size, improved neurological outcome | Pre- or early postischemia; continuous infusion or 1–20 mg/kg | Mixed animal results; failed large clinical trials in stroke [57] |
| Minocycline | Tetracycline | Anti-inflammatory (decreased microglial activation), anti-apoptotic (decreased caspase-3), decreased high-mobility group box 1, NF-κB | Middle cerebral artery occlusion-induced global ischemia in rat, mouse, and cat | Decreased infarct size, decreased apoptosis, improved long-term functional recovery | 30 min–24 h postischemia; 10–90 mg/kg | Excellent blood–brain barrier penetration; safe in humans but modest efficacy [58] |
| Memantine | N-methyl-D-aspartate receptor antagonist | Partial N-methyl-D-aspartate blockade limits excitotoxicity while preserving physiological signaling | Middle cerebral artery-induced hypertensive rat models | Decreased neuronal loss, improved cognitive outcome | Early postischemia; variable dosing | Better tolerated than dizocilpine; limited stroke-specific data [59] |
| Carvacrol | Natural monoterpenoid phenol | Antioxidant, anti-inflammatory, anti-apoptotic; inhibits transient receptor potential melastatin 7 channels | Ischemia and hypoxia models in rat and mice | Decreased oxidative stress, decreased neuroinflammation, decreased apoptosis | Pre- or early postischemia; variable doses | Emerging natural neuroprotectant, preclinical stage [60] |
| Tissue plasminogen activator | Thrombolytic | Converts plasminogen to plasmin, resulting in clot lysis | Thromboembolic stroke models | Restores cerebral blood flow, reduces infarct size, with hemorrhage risk | ≤3–4.5 h postischemia | Gold standard in clinics; often combined with neuroprotectants in animals [61] |
Herbal products as phytochemicals
Herbal products exhibit high variability in their medicinal value, particularly in antioxidant properties. This may be due to differences in species, the part of the plant used, climate conditions, and availability of these plants [62, 63]. Characterization and standardization of herbal products are based on various chemical constituents known as phytoconstituents. These phytoconstituents naturally occur in medicinal plants, leaves, seeds, vegetables, and roots and are responsible for multiple medicinal and pharmaceutical properties. They are classified into primary compounds, such as sugars and fats, and secondary compounds, such as alkaloids, tannins, terpenoids, phenols, and glycosides. These secondary compounds and other plant-derived chemical entities, such as Rheo discolor, are responsible for their medicinal value [64]. They have high contents of phenols (48.41 mg/1 g), flavonoids (43 mg/1 g), and tannins (44.93 mg/1 g). An earlier report found that phenols have strong antioxidant properties and reduce oxidative stress in the brain [65]. Flavonoids have been explored for their ability to prevent oxidative stress caused by ischemia–reperfusion. Tannins have also been reported to possess antioxidant and free-radical-scavenging activities [66]. These natural medicines are successful in preclinical studies because they are neuroprotective, ameliorating protein aggregation, oxidative stress, and neuroinflammation [67]. A comparison of the antioxidant content and strength of
Table 3. Comparison of the antioxidant content and strength of
| Herb | Antioxidant content | Relative strength |
|---|---|---|
| Withania somnifera (ashwagandha, roots/leaves) | Withanolides, flavonoids, and phenolics | Comparable to or slightly lower than |
| Curcuma longa (turmeric, rhizome) | Curcuminoids and phenolic acids | Stronger than |
| Ocimum sanctum (tulsi, leaves) | Eugenol, rosmarinic acid, and flavonoids | Comparable to or slightly higher than |
| Moringa oleifera (leaves) | Quercetin, chlorogenic acid, and vitamins | Much stronger than |
| Camellia sinensis (green tea, leaves) | Catechins | Much stronger than |
M. pruriens
Medicinal plants are gifts for fighting disease and death. Although many plants have been examined to date, only a handful have been studied thoroughly. The
Functional components of M. pruriens
Active components identified in M. pruriens
Its primary active compounds include levodopa, which is present at the highest level in its seeds, ranging from 0.2% to 7.30% of dry weight [72]. Its methanolic extracts yielded 3β-hydroxy-5α-cholanic acid acetate, 3,5,7,4-tetrahydroxy-6-methoxyflavone, and ethyl 2-amino-5-hydroxy-3,6,6-trimethyl heptonate. Other bioactive compounds, such as medicarpin and parvisoflavone B, have been identified as β-glucosidase inhibitors [73]. Ursolic acid and betulinic acid, which exhibit neuroprotective activities similar to those of levodopa [74], have also been detected. The medicinal properties of
Table 4. Phytochemical profile of
| Phytochemical | Compound | Plant parts | Major pharmacological relevance |
|---|---|---|---|
| Non-protein amino acids | Levodopa | Seeds, roots, leaves | Dopamine precursor; anti-Parkinson’s activity; neuroprotection; antioxidant effects [77] |
| Alkaloids | Mucunine, mucunadine, prurienine, nicotine | Seeds, leaves | Central nervous system activity; antimicrobial; contribution to neuropharmacology [78] |
| Indole amines | Serotonin, tryptamine | Pods, hairs, seeds | Neurotransmission; role in neuromodulation |
| Flavonoids | Quercetin, kaempferol, rutin | Leaves, seeds | Antioxidant, anti-inflammatory [79] |
| Phenolic compounds | Gallic acid, caffeic acid, ferulic acid | Seeds, leaves | Strong antioxidant activity; cytoprotective effects |
| Tannins | Condensed and hydrolyzable tannins | Seeds, leaves | Antimicrobial, antioxidant, protein-binding properties |
| Saponins | Triterpenoid saponins | Seeds | Immunomodulatory effects [77] |
| Sterols/triterpenes | β-Sitosterol, stigmasterol | Seeds, leaves | Anti-inflammatory |
| Proteins and peptides | Lectins, protease inhibitors | Seeds | Immune modulation [77] |
| Other bioactives | Amino acids, nucleosides, vitamins, phenolic derivatives | Seeds | Antioxidant, cytoprotective [80] |
Mechanism of action of the active components of M. pruriens
Levodopa reportedly enhances the ability to scavenge free radicals and reduce oxidative damage in cerebral ischemia [74]. It can cross the blood–brain barrier and restore dopaminergic tone. Flavonoids, polyphenols, and tannins scavenge nitric oxide, hydroxyl radicals, and superoxide. The levels of antioxidant enzymes, including CAT, SOD, and GPx, are increased, whereas MDA and lipid peroxidation are decreased. β-Sitosterol acts as a neuroprotective agent against cerebrovascular diseases by downregulating N-methyl-D-aspartate receptor gene expression and decreasing calcium influx, which prevents excitotoxic neuronal death [67]. β-Sitosterol is also involved in the suppression of ischemia-induced tau hyperphosphorylation and preservation of the integrity and stability of axons and microtubules.
Traditional uses of M. pruriens
The bioactive molecules of
Effects of M. pruriens in cerebral ischemia
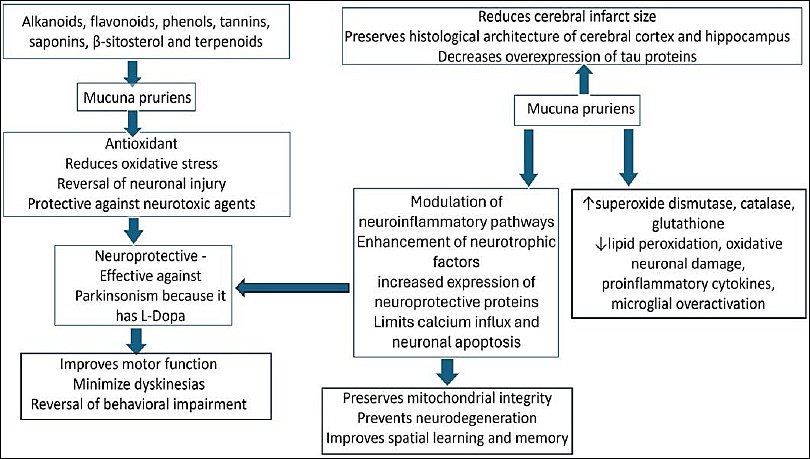
Figure 2. Schematic representation of the mechanism of action and neuroprotective role of
Challenges and limitations of the use of M. pruriens as neuroprotective agents
Translational barriers
The application of translational research and results from animal model studies to human clinical trials and experimental setups may be uncertain. Variability among individual subjects and efficacy across various species present significant challenges. There is a need for better guidelines to achieve significant therapeutic outcomes [23, 98, 99].
Regulatory guidelines
It has also been reported that unprocessed
Future implications
Although
CONCLUSION
The present review highlights that cerebral ischemia is a multifactorial neurological disorder characterized by oxidative stress, neuroinflammation, excitotoxicity, mitochondrial dysfunction, and apoptosis, ultimately leading to neuronal damage and functional impairment. Evidence from animal model studies consistently demonstrates that
From a practical perspective, the findings of this review suggest that
A major strength of this review lies in its comprehensive synthesis of animal model–based evidence, integrating biochemical, histopathological, and behavioral outcomes to provide a holistic understanding of the neuroprotective role of
However, several limitations must be acknowledged. The variability in experimental design, dosages, extraction methods, and treatment durations across studies limits the consistency and comparability of results. In addition, differences in animal species and models restrict the direct extrapolation of findings to human clinical conditions. The lack of standardized formulations and insufficient clinical trials further constrain the therapeutic application of
In conclusion,
DATA AVAILABILITY
The supplementary data can be made available from the corresponding author upon request.
AUTHORS’ CONTRIBUTIONS
VSN, KSRP, and SSN: Performed data acquisition and analysis. VSN, SSN, BVM, and SS: Wrote the manuscript. KSRP, SS, and MT: Manuscript review. All authors have read, reviewed, and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors thank the non-teaching staff members of their department for the help offered. The authors did not receive any funds for this study.
REFERENCES
- Zhu W, He X, Huang D, Jiang Y, Hong W, Ke S. Global and regional burden of ischemic stroke disease from 1990 to 2021: An age-period-cohort analysis. Transl Stroke Res 2025;16(5):1474-1485. [Google Scholar] | [Crossref]
- Hou S, Zhang Y, Xia Y, Liu Y, Deng X, Wang W. Global, regional, and national epidemiology of ischemic stroke from 1990 to 2021. Eur J Neurol 2024;31(12):e16481. [Google Scholar] | [Crossref]
- Neumar RW. Molecular mechanisms of ischemic neuronal injury. Ann Emerg Med 2000;36(5):483-506. [Google Scholar] | [Crossref]
- Ozbal S, Erbil G, Kocdor H, Tugyan K, Pekcetin C, Ozogul C. The effects of selenium against cerebral ischemia–reperfusion injury in rats. Neurosci Lett 2008;438(3):265-269. [Google Scholar] | [Crossref]
- Broughton BR, Reutens DC, Sobey CG. Apoptotic mechanisms after cerebral ischemia. Stroke 2009;40(5):e331-e339. [Google Scholar] | [Crossref]
- Malhotra K, Liebeskind DS. Collaterals in ischemic stroke. Brain Hemorrhages 2020;1(1):6-12. [Google Scholar] | [Crossref]
- Wu L, Xiong X, Wu X, Ye Y, Jian Z, Zhi Z. Targeting oxidative stress and inflammation to prevent ischemia–reperfusion injury. Front Mol Neurosci 2020;13:28. [Google Scholar] | [Crossref]
- Cui H, Kong Y, Zhang H. Oxidative stress, mitochondrial dysfunction, and aging. J Signal Transduct 2012;2012:646354. [Google Scholar] | [Crossref]
- Zengin G, Terzić M, Abul N, Gülçin İ, Koyuncu İ, Basarali MK. A multidimensional study for design functional foods: Chemical profiling, antioxidant potential, enzyme inhibition, and cytotoxic effects of Alkanna tubulosa extracts. Food Biosci 2025;60:104280. [Google Scholar] | [Crossref]
- Sims NR, Anderson MF, Hobbs LM, Kong JY, Phillips S, Powell JA. Impairment of brain mitochondrial function by hydrogen peroxide. Brain Res Mol Brain Res 2000;77(2):176-184. [Google Scholar] | [Crossref]
- Smith WS. Pathophysiology of focal cerebral ischemia: A therapeutic perspective. J Vasc Interv Radiol 2004;15(1):S3-S12. [Google Scholar] | [Crossref]
- Yapıcı I, Altay A, Ozturk Sarikaya SB, Korkmaz M, Atila A, Gulçin İ.
In vitro antioxidant and cytotoxic activities of extracts of endemic Tanacetum erzincanense together with phenolic content by LC-ESI-QTOF-MS. Chem Biodivers 2021;18(3):e2000812. [Google Scholar] | [Crossref] - Struneck A, Patocka J, Shapiro P, Atwood D. Aluminofluoride complexes: A useful tool in laboratory investigations, but a hidden danger for living organisms?. Washington: ACS Symposium Series; 2002. p. 271-282. [Google Scholar]
- Chirumari K, Reddy PK. Dose-dependent effects of fluoride on neurochemical milieu in the hippocampus and neocortex of rat brain. Fluoride 2007;40(2):101-110. [Google Scholar] | [Crossref]
- Hu X, Guo Z, Shi Z, Zhen P, Zhou M. Morphological changes in CA3 pyramidal neurons after transient global ischemia. Neuroreport 2025;36(14):856-863. [Google Scholar] | [Crossref]
- Li H, Xue X, Li L, Li Y, Wang Y, Huang T. Aluminum-induced synaptic plasticity impairment via PI3K-Akt-mTOR signaling pathway. Neurotox Res 2020;37(4):996-1008. [Google Scholar] | [Crossref]
- Li H, Xue X, Li Z, Pan B, Hao Y, Niu Q. Aluminiuium-induced synaptic plasticity injury via the PHF8-H3K9me2-BDNF signaling pathway. Chemosphere 2020;244:125445. [Google Scholar] | [Crossref]
- Yang L, Qian X, Jiang H, Xie C. Inhibition of SIRT1/PGC-1αaxis exacerbates fluorine and aluminium induced neurotoxicity via Drp1-dependent aggravated mitochondrial fission. Mol Neurobiol 2025;63(1):177. [Google Scholar] | [Crossref]
- Xin W, Pan Y, Wei W, Tatenhorst L, Graf I, Popa-Wagner A. Preconditioned extracellular vesicles from hypoxic microglia reduce poststroke AQP4 depolarization, disturbed cerebrospinal fluid flow, astrogliosis, and neuroinflammation. Theranostics 2023;13(12):4197-4216. [Google Scholar] | [Crossref]
- Jain M, Das S, Lu PPY, Virmani G, Soman S, Thumu SCR. SRF is required for maintenance of astrocytes in non-reactive state in the mammalian brain. eNeuro 2021;8(1). [Google Scholar] | [Crossref]
- Chandran P, Chandramohan K, Iyer K, Michael FM, Seppan P, Venkatachalam S. Beneficial effects of ethanolic extract of the medicinal herb
Mucuna pruriens against oxidative stress and inflammation might be limited in contusive spinal cord injury. Biomed Pharmacol J 2022;15(1):235-248. [Google Scholar] | [Crossref] - Bhosle PV, Wadher SJ. Evaluation of
Mucuna pruriens extract as a potential treatment for Huntington's disease: Antioxidant and anti-inflammatory mechanisms in rat models. J Neurosci Rural Pract 2025;16(4):524-532. [Google Scholar] | [Crossref] - Nayak VS, Kumar N, D'Souza AS, Nayak SS, Cheruku SP, Pai KSR. The effects of
Mucuna pruriens extract on histopathological and biochemical features in the rat model of ischemia. Neuroreport 2017;28(18):1195-1201. [Google Scholar] | [Crossref] - Dogra N, Nagpal D, Aeri V, Ahmad S, Pande Katare D. Evaluating the synergistic effect of Mucuna prurines extract and sesame oil against the Parkinson's disease zebrafish model:
In vivo /in silico approach. All Life 2021;14(1):1022-1042. [Google Scholar] | [Crossref] - Yadav SK, Prakash J, Chouhan S, Westfall S, Verma M, Singh TD. Comparison of the neuroprotective potential of
Mucuna pruriens seed extract with estrogen in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced PD mice model. Neurochem Int 2014;65:1-13. [Google Scholar] | [Crossref] - Manyam BV, Dhanasekaran M, Hare TA. Neuroprotective effects of the antiparkinson drug
Mucuna pruriens . Phytother Res 2004;18(9):706-712. [Google Scholar] | [Crossref] - Chukanova AS, Chukanova EI, Nadareishvili GG, Gulieva MS, Gusev EI. Patogeneticheskie aspekty formirovaniia ostroĭfokal'noĭishemii golovnogo mozga [Pathogenetic aspects of the development of acute focal cerebral ischemia. Zh Nevrol Psikhiatr Im S S Korsakova 2017;117(12 Pt 2):4-10. [Google Scholar] | [Crossref]
- Schaller BJ. Influence of age on stroke and preconditioning-induced ischemic tolerance in the brain. Exp Neurol 2007;205(1):9-19. [Google Scholar] | [Crossref]
- Quillinan N, Herson PS, Traystman RJ. Neuropathophysiology of brain injury. Anesthesiol Clin 2016;34(3):453-464. [Google Scholar] | [Crossref]
- Li Z, Li M, Fang Z, Wang H. Immunological mechanisms and therapeutic strategies in cerebral ischemia–reperfusion injury: From inflammatory response to neurorepair. Int J Mol Sci 2025;26(17):8336. [Google Scholar] | [Crossref]
- Mader MM, Heimann A, Kempski O, Wöbker G, Alessandri B. Multiparametric monitoring of early pathophysiological changes in a porcine model of sequential focal and global cerebral ischemia. World Neurosurg 2022;161:e473-e481. [Google Scholar] | [Crossref]
- Ister R, Pongrac M, DobrivojevićRadmilović M. Optimization of the longa middle cerebral artery occlusion method for complete reperfusion. J Vis Exp 2024;213. [Google Scholar] | [Crossref]
- Ahad MA, Kumaran KR, Ning T, Mansor NI, Effendy MA, Damodaran T. Insights into the neuropathology of cerebral ischemia and its mechanisms. Rev Neurosci 2020;31(5):521-538. [Google Scholar] | [Crossref]
- Pan J, Konstas AA, Bateman B, Ortolano GA, Pile-Spellman J. Reperfusion injury following cerebral ischemia: Pathophysiology, MR imaging, and potential therapies. Neuroradiology 2007;49(2):93-102. [Google Scholar] | [Crossref]
- Kahl A, Stepanova A, Konrad C, Anderson C, Manfredi G, Zhou P. Critical role of flavin and glutathione in complex I-mediated bioenergetic failure in brain ischemia/reperfusion injury. Stroke 2018;49(5):1223-1231. [Google Scholar] | [Crossref]
- Luo T, Park Y, Sun X, Liu C, Hu B. Protein misfolding, aggregation, and autophagy after brain ischemia. Transl Stroke Res 2013;4(6):581-588. [Google Scholar] | [Crossref]
- Zhang Y, Liu L, Hou X, Zhang Z, Zhou X, Gao W. Role of autophagy mediated by AMPK/DDiT4/mTOR axis in HT22 cells under oxygen and glucose deprivation/reoxygenation. ACS Omega 2023;8(10):9221-9229. [Google Scholar] | [Crossref]
- Wang L, Zhang X, Xiong X, Zhu H, Chen R, Zhang S. Nrf2 regulates oxidative stress and its role in cerebral ischemic stroke. Antioxidants (Basel) 2022;11(12):2377. [Google Scholar] | [Crossref]
- Li Y, Jiang J, Zhuo Y, Li J, Li Y, Xia Y. IGF2BP1 exacerbates neuroinflammation and cerebral ischemia/reperfusion injury by regulating neuronal ferroptosis and microglial polarization. Biochim Biophys Acta Mol Basis Dis 2025;1871(6):167877. [Google Scholar] | [Crossref]
- Gupta YK, Briyal S. Animal models of cerebral ischemia for evaluation of drugs. Indian J Physiol Pharmacol 2004;48(4):379-394. [Google Scholar] | [Crossref]
- Li T, Xiam J, Dong Y, Chen J, Ren M. Controlled intracranial pressure elevation via cerebrospinal fluid infusion: A novel hemostatic hypothesis for hematoma expansion in traumatic brain contusion. Medical Hypotheses 2026;206:111837. [Google Scholar] | [Crossref]
- Basilio AV, Zeng D, Pichay LA, Ateshian GA, Xu P, Maas SA. Simulating cerebral edema and ischemia after traumatic acute subdural hematoma using triphasic swelling biomechanics. Ann Biomed Eng 2024;52(10):2818-2830. [Google Scholar] | [Crossref]
- Donnelly J, Czosnyka M, Harland S, Varsos GV, Cardim D, Robba C. Increased ICP and its cerebral haemodynamic sequelae. Acta Neurochir Suppl 2018;126:47-50. [Google Scholar] | [Crossref]
- Lafrenaye AD, Krahe TE, Povlishock JT. Moderately elevated intracranial pressure after diffuse traumatic brain injury is associated with exacerbated neuronal pathology and behavioral morbidity in the rat. J Cereb Blood Flow Metab 2014;34(10):1628-1636. [Google Scholar] | [Crossref]
- Verma R, Bisen P.
Mucuna pruriens - a wonder herb for degenerative disorders. In:An introduction to medicinal herbs. Chapter 8. Nova Science Publisher 2021:237-258. [Google Scholar] | [Crossref] - Jin X, Jing W, Fengxia Y, Zhang Z, Fengjun L, Jing S. Effect of intracranial hypertension on cerebral hemorrhage induced autonomic nerve imbalance. Neural Reg Research 2007;2(3):175-178. [Google Scholar] | [Crossref]
- Abdou H, Treffalls R, Jodlowski G, Elansary N, Ptak T, Walker PF. The influence of hemorrhagic shock on brain perfusion in a swine model of raised intracranial pressure. Eur J Trauma Emerg Surg 2025;51(1):137. [Google Scholar] | [Crossref]
- Hamilton DR, Sargsyan AE, Melton SL, Garcia KM, Oddo B, Kwon DS. Sonography for determining the optic nerve sheath diameter with increasing intracranial pressure in a porcine model. J Ultrasound Med 2011;30(5):651-659. [Google Scholar] | [Crossref]
- Calloni RL, Winkler BC, Ricci G, Poletto MG, Homero WM, Serafini EP. Transient middle cerebral artery occlusion in rats as an experimental model of brain ischemia. Acta Cir Bras 2010;25(5):428-433. [Google Scholar] | [Crossref]
- Vidale S, Consoli A, Arnaboldi M, Consoli D. Postischemic inflammation in acute stroke. J Clin Neurol 2017;13(1):1-9. [Google Scholar] | [Crossref]
- Gerriets T, Stolz E, Walberer M, Müller C, Rottger C, Kluge A. Complications and pitfalls in rat stroke models for middle cerebral artery occlusion: A comparison between the suture and the macrosphere model using magnetic resonance angiography. Stroke 2004;35(10):2372-2377. [Google Scholar] | [Crossref]
- Nagel S, Papadakis M, Chen R, Hoyte LC, Brooks KJ, Gallichan D. Neuroprotection by dimethyloxalylglycine following permanent and transient focal cerebral ischemia in rats. J Cereb Blood Flow Metab 2011;31(1):132-143. [Google Scholar] | [Crossref]
- Nour M, Scalzo F, Liebeskind DS. Ischemia-reperfusion injury in stroke. Intervent Neurol 2013;1(3-4):185-199. [Google Scholar] | [Crossref]
- McCulloch J, Ozyurt E, Park CK, Nehls DG, Teasdale GM, Graham DI, Baethmann A, Kempski O, Schürer L. Glutamate receptor antagonists in experimental focal cerebral ischaemia. Vienna: Springer; 1993. p. 57. [Google Scholar]
- Zhao LQ, Parikh A, Xiong YX, Ye QY, Ying-Guo, Zhou XF. Neuroprotection of oral edaravone on middle cerebral artery occlusion in rats. Neurotox Res 2022;40:995-1006. [Google Scholar] | [Crossref]
- Rahmati-Dehkordi F, Khanifar H, Zare-Hoseinabadi A, Dadgostar E, Jafarpour H, Aschner M. Potential of edaravone dexborneol in the treatment of cerebral ischemia: Focus on cell death-related signaling pathways. Mol Biol Rep 2024;51:1007. [Google Scholar] | [Crossref]
- Horn J, de Haan RJ, Vermeulen M, Luiten PG, Limburg M. Nimodipine in animal model experiments of focal cerebral ischemia: A systematic review. Stroke 2001;32(10):2433-2438. [Google Scholar] | [Crossref]
- Tang XN, Wang Q, Koike MA, Cheng D, Goris ML, Blankenberg FG. Monitoring the protective effects of minocycline treatment with radiolabeled annexin V in an experimental model of focal cerebral ischemia. J Nucl Med 2007;48(11):1822-1828. [Google Scholar] | [Crossref]
- Song X, Jensen MØ, Jogini V, Stein RA, Lee CH, Mchaourab HS. Mechanism of NMDA receptor channel block by MK-801 and memantine. Nature 2018;556:515-519. [Google Scholar] | [Crossref]
- Zhang XY, Ho HL, Feng ZP, Sun HS. Neuroprotective effects of carvacrol in cerebral ischemia and hypoxia. International Journal of Drug Discovery and Pharmacology 2026;5(1):100003. [Google Scholar] | [Crossref]
- Thaysen M, Westi E, Clarkson AN, Wellendorph P, Kristensen M. Rodent ischemic stroke models and their relevance in preclinical research. Neuroprotection 2024;2(4):296-309. [Google Scholar] | [Crossref]
- Rodriguez-Amaya DB. Food carotenoids: Analysis, composition and alterations during storage and processing of foods. Forum Nutr 2003;56:35-37. [Google Scholar] | [Crossref]
- Yadav NP, Dixit VK. Recent approaches in herbal drug standardization. Int J Integr Biol 2008;2(3):195-203. [Google Scholar] | [Crossref]
- Parivuguna V, Gnanaprabhal R, Dhanabalan R, Doss A. Antimicrobial properties and phytochemical constituents of Rheo discolour Hance. Ethnobotanical Leaflets 2008;12:841-845. [Google Scholar] | [Crossref]
- Beauchamp GK, Keast RS, Morel D, Lin J, Pika J, Han Q. Phytochemistry: Ibuprofen-like activity in extra-virgin olive oil. Nature 2005;437(7055):45-46. [Google Scholar] | [Crossref]
- Smeriglio A, Barreca D, Bellocco E, Trombetta D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br J Pharmacol 2017;174(11):1244-1262. [Google Scholar] | [Crossref]
- Das S, Rajeswari VD, Venkatraman G, Ramanathan G. Phytochemicals in Parkinson's disease: A pathway to neuroprotection and personalized medicine. Cell Biochem Biophys. [Google Scholar] | [Crossref]
- Alam N, Hossain M, Mottalib MA, Sulaiman SA, Gan SH, Khalil MI. Methanolic extracts of Withania somnifera leaves, fruits and roots possess antioxidant properties and antibacterial activities. BMC Complement Altern Med 2012;12:175. [Google Scholar] | [Crossref]
- Narayanasamy A, Kanagaraja A, Thirumavalavan M, Sakthivelu M, Pachaiappan R. Evaluation of antioxidant activities of bioactive peptides extracted from Curcuma longa and Curcuma caesia from South-eastern and North-Eastern India. Probiotics Antimicro Prot 2026. [Google Scholar] | [Crossref]
- Osuntokun OS, Olayiwola G, Oriare AK, Oyedokun SO, Abayomi TA, Tokunbo OS.
Mucuna pruriens seed protects the hippocampal neurons and abrogates seizure indices in chemically-convulsed mice: Evidence of the Nrf2 expression defense pathway. J Chem Neuroanat 2022;123:102115. [Google Scholar] | [Crossref] - Lampariello LR, Cortelazzo A, Guerranti R, Sticozzi C, Valacchi G. The magic velvet bean of
Mucuna pruriens . J Tradit Complement Med 2012;2(4):331-339. [Google Scholar] | [Crossref] - Hammoud F, Ismail A, Zaher R, El Majzoub R, Abou-Abbas L.
Mucuna pruriens treatment for Parkinson's disease: A systematic review of clinical trials. Parkinson's Dis 2025;2025:1319419. [Google Scholar] | [Crossref] - Dendup T, Prachyawarakorn V, Pansanit A, Mahidol C, Ruchirawat S, Kittakoop P. α-Glucosidase inhibitory activities of isoflavanones, isoflavones, and pterocarpans from
Mucuna pruriens . Planta Med 2014;80(7):604-608. [Google Scholar] | [Crossref] - Rai SN, Chaturvedi VK, Singh P, Singh BK, Singh MP.
Mucuna pruriens in Parkinson's and in some other diseases: Recent advancement and future prospective. 3 Biotech 2020;10(12):522. [Google Scholar] | [Crossref] - Aslam S, Rafiq A, Ahmad M, Naqvi SAR, Al-Huqail AA, Zia-Ul-Haq M, Al-Huqail AA, Riaz M, Farooq Gohar U. Cowhage. Cham: Springer; 2023. p. 145-169. [Google Scholar]
- Pathania R, Chawla P, Khan H, Kaushik R, Khan MA. An assessment of potential nutritive and medicinal properties of
Mucuna pruriens : A natural food legume. 3 Biotech 2020;10(6):261. [Google Scholar] | [Crossref] - Deli M, Nguimbou RM, Djantou EB, Tatsadjieu Ngoune L, Njintang Yanou N, Murthy HN, Paek KY. Bioactive compounds of velvet bean (
Mucuna pruriens L.). seeds. Cham: Springer; 2021. [Google Scholar] - Yadav M, Upadhyay P, Purohit P, Pandey B, Shah H. Phytochemistry and pharmacological activity of
Mucuna pruriens : A review. International Journal of Green Pharmacy 2017;11:69-73. [Google Scholar] | [Crossref] - Jimoh MA, Idris OA, Jimoh MO. Cytotoxicity, phytochemical, antiparasitic screening, and antioxidant activities of
Mucuna pruriens (Fabaceae). Plants 2020;9:1249. [Google Scholar] | [Crossref] - Sruthi D, Jayabaskaran C. Performance of different solvents and extraction methods on therapeutic potential of
Mucuna pruriens (L.). DC. and chemical profiling of screened extract with chromatography-mass spectrometry approach. Curr Res Cmpl Alt Med 2024;8:237. [Google Scholar] | [Crossref] - Mata-Bermudez A, Diaz-Ruiz A, Silva-García LR, Gines-Francisco EM, Noriega-Navarro R, Rios C.
Mucuna pruriens , a possible treatment for depressive disorders. Neurol Int 2024;16(6):1509-1527. [Google Scholar] | [Crossref] - Zaigham SB, Paeng DG. Effects of
Mucuna pruriens (L.). DC. and levodopa in improving Parkinson's disease in rotenone intoxicated mice. Curr Issues Mol Biol 2024;46(8):9234-9244. [Google Scholar] | [Crossref] - Adi YK, Widayanti R, Pangestiningsih TW. n-Propanol extract of boiled and fermented koro benguk (
Mucuna pruriens seed) shows a neuroprotective effect in paraquat dichloride-induced Parkinson's disease rat model. Vet World 2018;11(9):1250-1254. [Google Scholar] | [Crossref] - Osuntokun OS, Olayiwola G, Oriare AK, Odeniran HT, Ayoka AO. The activities of the central nervous system following ethyl acetate extract of
Mucuna pruriens seed administration in male BALB/c mice. Nig J Neurosci 2021;12(3):92-99. [Google Scholar] | [Crossref] - Concessao P, Bairy LK, Raghavendra AP. Protective effect of
Mucuna pruriens against arsenic-induced liver and kidney dysfunction and neurobehavioral alterations in rats. Vet World 2020;13(8):1555-1566. [Google Scholar] | [Crossref] - Mata-Bermudez A, Trejo-Chávez R, Martínez-Vargas M, Pérez-Arredondo A, de Los Ángeles, Martínez-Cardenas M. Effect of
Mucuna pruriens seed extract on depression-like behavior derived from mild traumatic brain injury in rats. Biomedicine (Taipei) 2024;14(3):23-30. [Google Scholar] | [Crossref] - Sathiyanarayanan L, Arulmozhi S.
Mucuna pruriens : A comprehensive review. Pharmacogn Rev 2007;1(1):157-162. [Google Scholar] | [Crossref] - Bhosle S, Bagali S, Parvatikar PP, Das KK. Effect of bioactive compounds of
Mucuna pruriens on proteins of Wnt/βcatenin pathway in pulmonary hypertension byin silico approach. In Silico Pharmacol 2024;12(2):110. [Google Scholar] | [Crossref] - Guerranti R, Ogueli IG, Bertocci E, Muzzi C, Aguiyi JC, Cianti R. Proteomic analysis of the pathophysiological process involved in the antisnake venom effect of
Mucuna pruriens extract. Proteomics 2008;8(2):402-412. [Google Scholar] | [Crossref] - Misra L, Wagner H. Extraction of bioactive principles from
Mucuna pruriens seeds. Indian J Biochem Biophys 2007;44(1):56-60. [Google Scholar] | [Crossref] - Arif M, Alam M, Malhi SM, Azeem Z, Sherwani B, Asghar MN.
In vivo evaluation of medicinal effects of Myristica fragrans, Cinnamomum zeylanicum,Mucuna pruriens on male murine fertility. Pak J Pharm Sci 2024;37(3):601-611. [Google Scholar] | [Crossref] - Concessao PL, Bairy KL, Raghavendra AP. Ameliorating effect of
Mucuna pruriens seed extract on sodium arsenite-induced testicular toxicity and hepato-renal histopathology in rats. Vet World 2023;16(1):82-93. [Google Scholar] | [Crossref] - Wattanawiggan R, Chansakaow S, Jantrawut P, Panraksa P, Jiaranaikulwanitch J, Udomsom S. Design and optimization of 3D-printed tablets containing Mucuna extracts for erectile dysfunction management: A DoE-guided study. Plants (Basel) 2024;13(16):2294. [Google Scholar] | [Crossref]
- Kumbhare SD, Ukey SS, Gogle DP. Antioxidant activity of Flemingia praecox and
Mucuna pruriens and their implications for male fertility improvement. Sci Rep 2023;13(1):19360. [Google Scholar] | [Crossref] - Samrid R, Taoto C, Wu A, Sawatpanich T, Phunchago N, Uabundit N. Protective effect of
Mucuna pruriens (L.). DC. var. pruriens seed extract on apoptotic germ cells in ethanolic male rats. Braz J Biol 2023;83:e272629. [Google Scholar] | [Crossref] - Beta T, Nam S, Dexter JE, Sapirstein HD. Phenolic content and antioxidant activity of pearled wheat and roller-milled fractions. Cereal Chem 2005;82(4):390-393. [Google Scholar] | [Crossref]
- Alabi AO, Ogunjimi LO, Murtala AA, Kasumu EO, Oyinloye EO, Shofoyeke AM. Sub-acute toxicity, antinociceptive and anti-inflammatory effects of
Mucuna pruriens L. leaves in experimental rodents. J Ethnopharmacol 2024;333:118489. [Google Scholar] | [Crossref] - Parvatikar PP, Patil SM, Patil BS, Reddy RC, Bagoji I, Kotennavar MS. Effect of
Mucuna pruriens on brain NMDA receptor and tau protein gene expression in cerebral ischemic rats. Front Physiol 2023;14:1092032. [Google Scholar] | [Crossref] - Nayak VS, Pai KSR, Nayak SS, Kumar N, Bangera H. Effect of
Mucuna pruriens (Linn.). on global cerebral ischemia-induced motor incoordination. Trop J Pharma Res 2021;20(6):1193-1198. [Google Scholar] | [Crossref] - Oyinloye OE, Murtala AA, Oladoja FA, Okunye OL, Aderinola AA, Kasumu EO. Evaluation of phytochemical constituents, total phenolic contents and antioxidant activities of
Mucuna pruriens fractions leaves. J Phytomed Ther 2023;22(1):1017-1034. [Google Scholar] | [Crossref] - Kumar DS, Muthu AK, Smith AA, Manavalan R.
In vivo antioxidant and lipid peroxidation effect of various extracts of whole plant ofMucuna pruriens (Linn) in rat fed with high-fat diet. Asian J Chem 2011;23(2):823-825. [Google Scholar] | [Crossref] - Krishna RG, Sundararajan R. Screening of antioxidant activity of
Mucuna pruriens byin vivo model. Int J Res Pharm Sci 2019;10(1):523-530. [Google Scholar] | [Crossref] - Kumar N, Singh SK, Lal RK, Dhawan SS. An insight into dietetic and nutraceutical properties of underutilized legume:
Mucuna pruriens (L.) DC. J Food Comp Anal 2024;129:106095. [Google Scholar] | [Crossref] - Panova AS, Dergachev DS, Subotyalov MA, Dergachev VD. Review of
Mucuna pruriens L. therapeutic potential for Parkinson's disease. Meditsinskiy Sovet 2020;8:82-87. [Google Scholar] | [Crossref] - Sato K, Hida A, Niimi Y, Iwata A, Iwatsubo T. Survey on the current advertising and sales of
Mucuna pruriens in consumer-to-consumer internet trading in Japan. Yakugaku Zasshi 2023;143(12):1057-1067. [Google Scholar] | [Crossref]