


ABSTRACT
Background and Aim: Legume-based green concentrate (GC) is a promising, low-cost feeding strategy for dairy systems; however, its high rumen-degradable protein (RDP) fraction limits nitrogen utilization efficiency. This study evaluated the effect of
Materials and Methods: The experiment comprised integrated
Results:
Conclusion: Enrichment of GC with
Keywords: dairy nutrition,
INTRODUCTION
Average milk yield in Indonesia remains low at approximately 9–10 L/cow/day [1]. Beyond environmental constraints associated with tropical climates [2], milk production is strongly determined by feed quality. Nutritional inadequacies—particularly protein and energy—depress milk yield [3], whereas mineral imbalances (e.g., calcium) may precipitate hypocalcemia manifested as muscular stiffness or paresis [4]. Furthermore, early lactation is frequently characterized by negative energy balance (NEB), which heightens metabolic stress and compromises performance [5].
Concentrate feeds commonly offered to dairy cows are commercial formulations that are relatively costly and sporadically available. Consequently, farmers often limit concentrate provision, with downstream impacts on milk output. Developing locally available, accessible, and cost-effective alternatives is therefore a pressing need. Legume-based green concentrate (GC) represents a promising option because its protein sources derive from legumes such as
However, a key limitation of GC is its high proportion of rumen-degradable protein (RDP), which predisposes nitrogen losses in the rumen and constrains the post-ruminal supply of amino acids [7]. To address this limitation, protein protection technologies—namely rumen-protected agents (RPA), have been developed to increase rumen-undegradable protein (RUP) and improve intestinal amino acid delivery [9]. Among encapsulating materials, polysaccharides such as chitosan can form ionic gel networks in the rumen, thereby retarding proteolysis and improving nutrient utilization [10]. Chitosan has also been shown to enhance daily weight gain and feed efficiency in broilers [11].
Despite its efficacy, chitosan extraction from crab shells and other sources is technically demanding and costly for smallholder farmers. Kappa-carrageenan, a sulfated polysaccharide found predominantly in the seaweed
To date, the enrichment of GC with
Therefore, this study aimed to evaluate the effect of legume-based GC enriched with
MATERIALS AND METHODS
Ethical approval
All experimental procedures were conducted in accordance with institutional animal welfare guidelines. Ethical approval was obtained from the Ethics Committee on the Use of Research and Learning Animals, Faculty of Animal Science, Hasanuddin University, Indonesia (Approval No. 030/UN4.12/EC/VIII/2025). The dairy cows used in this study were clinically healthy and free from mastitis. Routine mastitis monitoring was conducted every 14 days at the research site, and post-milking teat dipping was consistently implemented as a preventive measure. Prior to animal allocation, a screening process was undertaken to ensure homogeneity in physiological and production characteristics. Randomization of experimental animals was then performed manually using a lottery procedure to ensure unbiased group assignment.
Study period and location
The experiment was conducted from August 8 to October 7, 2025 in Lebang Village, Cendana District, Enrekang Regency, South Sulawesi, Indonesia. The research location was 3°29′12.12″ South Latitude and 119°51′40.319″ East Longitude, at an altitude of 47 m above sea level (map coordinates). The study was done under real farm conditions. The average temperature in the cage ranged from 24°C (morning) to 34°C (afternoon), relative humidity was 85% (morning) and 52% (afternoon), and the cage received approximately 13 h of natural light (calculated during the research period). However, sample testing was conducted in the laboratory as an indicator of inter-treatment assessment after implementation in dairy cows to ensure the accuracy and validity of livestock performance evaluation results. The milk quality was analyzed at the Dairy Production Laboratory, Faculty of Animal Science, Hasanuddin University, Makassar. Quality of the diet was analyzed at the Dairy Animal Nutrition Laboratory, IPB University, Bogor, West Java.
Study design
This study was divided into two stages, including
All treatments were subjected to
The treatments consisted of CON, legume-based GC without enrichment (GC), and GC enriched with 10%
Diet formulation and feeding management
All diets were formulated to comply with national standards for lactating dairy cows concentrates, ensuring a minimum of 18% crude protein (CP) and 70% total digestible nutrients (Table 1).
Table 1. Composition of diets under treatments.
| Feedstuffs (%) | CON | GC |
|---|---|---|
| Soybean meal | 24 | 13 |
| Rice bran | 38 | 36 |
| Molasses | 2 | 2 |
| Corn Meal | 36 | 29 |
| Indigofera leaves meal | 0 | 5 |
| Gliricidia leaves meal | 0 | 15 |
| Total | 100 | 100 |
CON = Commercial concentrate, GC = Green concentrate.
Legume components, Gliricidia and Indigofera leaves, were harvested, dehydrated at 70°C for 7 h, and subsequently ground into meal using a disc mill. The same procedure was applied to
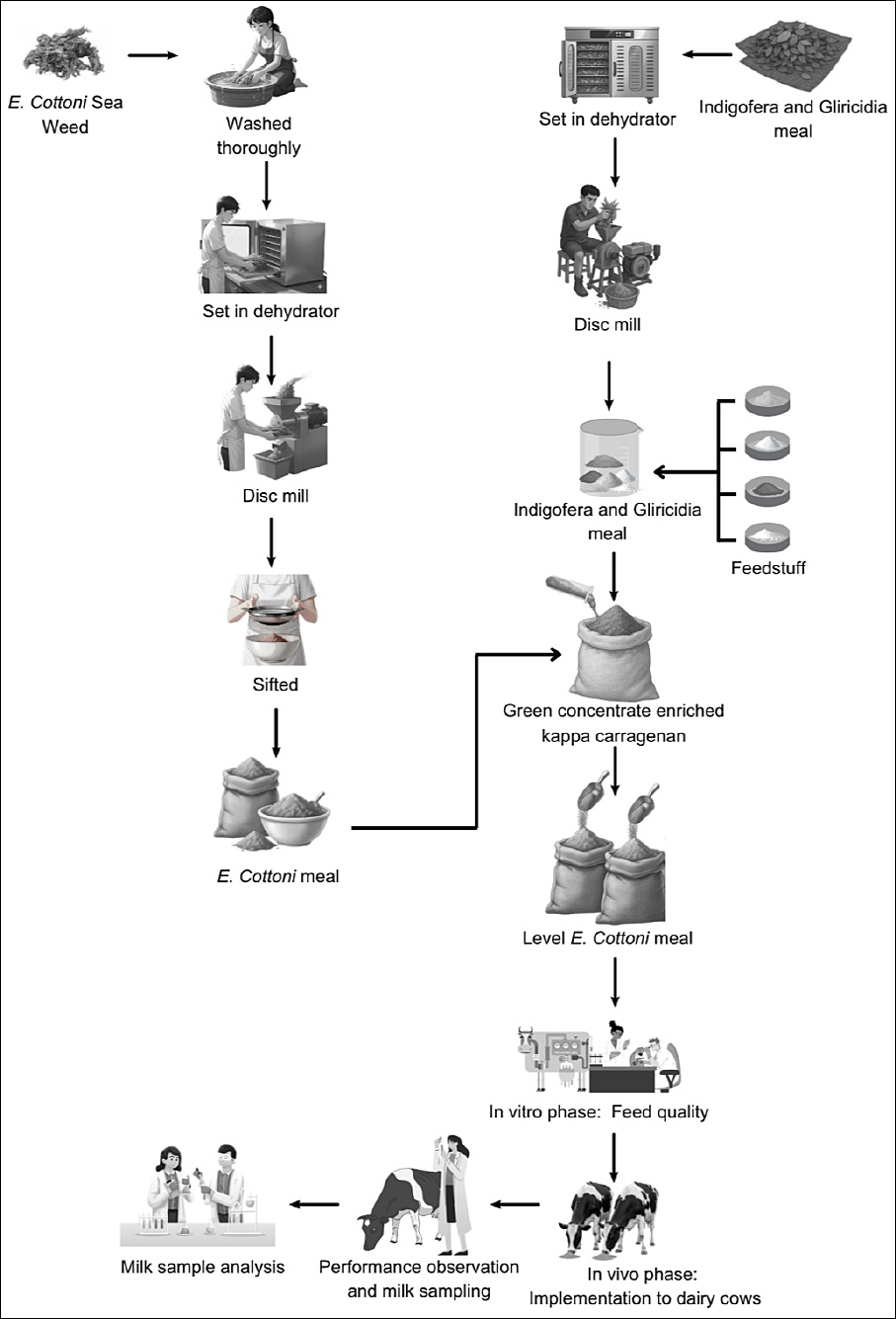
Figure 1. The diagram summarizes the experimental workflow, including diet formulation,
Table 2. Nutritional value of commercial concentrate and
| Nutritional composition (%) | Commercial concentrate | |
|---|---|---|
| CP | 18.79 | 6.31 |
| CF | 5.93 | 0.09 |
| CFi | 5.95 | 7.86 |
| Ash | 4.33 | 13.70 |
| Kappa-carrageenan | – | 60 |
CP = Crude protein, CF = Crude fat, CFi = Crude fiber, Nutritional values of
Seaweed
The commercial concentrate (Sipatuo®, CV Sipatuo Farm, Enrekang, Indonesia) used as a reference treatment was sourced from CV Sipatuo Farm; nutritional values refer to Table 2, a widely utilized brand among local dairy producers.
Cows were fed at 3% of body weight on a dry matter basis, with a concentrate-to-forage ratio of 30:70, using elephant grass (
Chemical composition analysis of the diet
Feed samples were analyzed for proximate composition following Association of Official Analytical Chemists (AOAC) protocols: CP; AOAC 976.05), crude fat (CF; AOAC 989.04), crude fiber (CFi; AOAC 962.09), nitrogen-free extract (NFE; AOAC 2003.05), and ash content (AOAC 942.05). Gross energy (GE) was determined using a bomb calorimeter (Model XRY-1A+, China, calibrated with benzoic acid standard), which measures calorific value based on the heat released during complete combustion of the sample in an oxygen-rich environment [15].
In vitro rumen fermentation and digestibility analysis
Rumen fluid was obtained from HF dairy cows kept in the field laboratory. The cows used had been fistulated to facilitate rumen fluid collection. Rumen fluid collection was carried out in the morning to ensure that the microenvironment was more representative. Dairy cows were fed a 70:30 forage:concentrate diet and strained through cheesecloth under CO2 flushing.
VFAs are produced during microbial fermentation of carbohydrates and proteins in the rumen. Major VFAs include acetate, propionate, isopropionate, butyrate, isobutyrate, valerate, and isovalerate. VFAs concentrations were analyzed using gas chromatography, based on differential absorption and partitioning across stationary and mobile phases. Separation resulted in distinct peaks on the chromatogram. Sample concentrations were quantified by comparing peak areas to those of known reference standards.
Digestibility was calculated using the Tilley and Terry two-stage technique [16]:
%IV digestibility = (initial weight (g) − (residual weight (g) − blank correction)) / (initial weight (g)) × 100%
The RDP and RUP levels, as well as rumen fluid characteristics
The gas production was measured every 2 h using a 10 mL syringe with a 0.1 mm needle injected into the rubber part of the sealed bottle during 24 h
Animal performance evaluation
Milk yield and dry matter (DM) intake were monitored over a 60-day period. Dry matter intake (DMI) is determined by subtracting the daily feed residue from the amount of feed offered (DM). The feed residue is weighed each day to provide the actual DMI value. FCE was calculated as the ratio of milk yield to DM intake (milk yield ÷ DM intake).
BCS was assessed on days 0, 30, and 60 of the trial. BCS was used as an indicator of cow health and was determined through visual inspection and palpation. BCS was evaluated following Edmonson
Economic analysis
Feed input costs (USD/kg DM) were calculated using prevailing market prices for each concentrate component. The GC cost was determined based on the current market price of its raw ingredients. Although forage was home-grown, costs were estimated based on fertilizer, labor, and transportation expenses.
Daily feed cost (USD/head/day) was computed by multiplying feed price (USD/kg DM) by feed intake (kg DM/head/day). Income from daily milk yield (USD/head/day) was calculated as milk yield (kg/day) × milk price (USD/kg). Daily profit (USD/head/day) was obtained by subtracting daily feed cost from daily milk income:
Daily profit = milk income − feed cost
Daily profit = (milk yield × milk price) − (DMI concentrate × concentrate price + DMI forage × forage price)
Milk quality assessment
On day 60, composite milk samples were collected from each cow for laboratory analysis. Approximately 100 mL of milk was obtained per animal, immediately placed in sterile containers, and stored in a cooler box to maintain sample integrity during transport to the laboratory. During transportation, the cooler box temperature remained stable at 4°C for a duration of 5 h.
Subsequent analyses included crude fat, protein, lactose, solids-not-fat (SNF), inorganic salt, and density, assessed using an automatic milk analyzer (Infitek MA-H3, China). The analyzer was calibrated daily using standard milk samples. Milk samples (50 mL) were homogenized at 3,000 rpm for 30 s and analyzed under controlled laboratory conditions (18–24°C). Two repetitions were carried out on the same milk sample, and the results used were the average of these tests.
Milk curd and whey percentages were measured by heating 1 kg of fresh milk using the low-temperature, long-time method. Then, 4 mL of papain enzyme from papaya latex solution was added. After separation, curd and whey were weighed and calculated following Sutomo
%Curd = (curd weight) / (milk initial weight (g)) × 100%
%Whey = (whey weight) / (milk initial weight (g)) × 100%
Statistical analysis
Data were analyzed using one-way analysis of variance in SPSS version 27.0 (IBM Corp., Armonk, NY, USA). The model used was:
Yij = μ + τi + εij
Where Yij is the observed value, μ is the overall mean, τi is the treatment effect, and εij is the residual error. Where significant differences were found, Duncan’s multiple range test was used for post hoc comparison (p < 0.05). Data normality was tested using the Shapiro–Wilk test, and homogeneity was assessed using Levene’s test. Pearson correlation analysis was conducted to evaluate the relationship between the inclusion level of
RESULTS
Chemical analysis, rumen protein, gas production, rumen fermentation, and nutrient digestibility of the diets
Statistical analysis (Table 3) revealed that enrichment of GC with microencapsulated
Table 3. Chemical analysis, rumen protein, gas production, rumen fermentation, and nutrient digestibility of GC fortified with
| Parameters | GC | GCC1 | GCC2 | GCC3 | GCC4 | SEM | p-value |
|---|---|---|---|---|---|---|---|
| Chemical analysis (%) | |||||||
| CP | 20.10 ± 0.82 | 19.13 ± 0.60 | 18.87 ± 0.72 | 18.96 ± 0.35 | 18.65 ± 0.72 | 0.19 | 0.152 |
| CF | 5.23 ± 0.12ᵇ | 4.93 ± 0.33ᵇ | 4.47 ± 0.19ᵃᵇ | 4.42 ± 0.49ᵃᵇ | 3.81 ± 0.71ᵃ | 0.16 | 0.020 |
| CFi | 5.30 ± 0.47ᵃ | 6.49 ± 0.25ᵇ | 6.45 ± 0.19ᵇ | 6.51 ± 0.34ᵇ | 6.68 ± 0.31ᵇ | 0.15 | 0.003 |
| Rumen protein utilization (%) | |||||||
| RDP | 72.18 ± 1.25ᶜ | 69.53 ± 0.52ᵇ | 69.40 ± 0.68ᵇ | 66.70 ± 1.15ᵃ | 65.24 ± 1.17ᵃ | 0.68 | 0.000 |
| RUP | 59.56 ± 0.85ᵃ | 60.43 ± 1.18ᵃᵇ | 60.57 ± 1.12ᵃᵇ | 62.42 ± 1.40ᵇᶜ | 64.17 ± 1.09ᶜ | 0.50 | 0.004 |
| Gas production | |||||||
| Methane gas (% of total gas) | 11.09 ± 0.13ᶜ | 10.86 ± 0.62ᵇᶜ | 10.24 ± 0.46ᵃᵇ | 9.85 ± 0.31ᵃ | 9.76 ± 0.41ᵃ | 0.16 | 0.011 |
| Total gas (mL/g) | 57.00 ± 2.00ᶜ | 56.66 ± 1.15ᶜ | 55.00 ± 3.46ᵇᶜ | 52.66 ± 1.15ᵃᵇ | 50.00 ± 1.00ᵃ | 0.82 | 0.007 |
| Rumen fermentation (mmol/L) | |||||||
| Acetate | 50.57 ± 10.69ᵃ | 52.51 ± 21.65ᵃ | 53.68 ± 20.78ᵃ | 32.07 ± 10.95ᵃ | 83.09 ± 12.04ᵇ | 5.59 | 0.036 |
| Propionate | 15.21 ± 4.19ᵃ | 28.27 ± 6.27ᵇ | 18.45 ± 4.19ᵃ | 11.68 ± 3.61ᵃ | 38.28 ± 3.41ᶜ | 2.76 | 0.000 |
| Isobutyrate | 0.43 ± 0.00ᵃ | 0.76 ± 0.14ᵃ | 0.57 ± 0.14ᵃ | 0.67 ± 0.37ᵃ | 1.26 ± 0.24ᵇ | 0.89 | 0.009 |
| Butyrate | 4.18 ± 0.03ᵃ | 7.28 ± 1.15ᵇ | 5.28 ± 0.98ᵃᵇ | 6.17 ± 1.91ᵃᵇ | 10.16 ± 1.52ᶜ | 0.61 | 0.002 |
| Isovalerate | 0.50 ± 0.01ᵃ | 0.88 ± 0.09ᵃ | 0.66 ± 0.16ᵃ | 0.81 ± 0.40ᵃ | 1.47 ± 0.32ᵇ | 0.10 | 0.006 |
| Valerate | 0.31 ± 0.01ᵃ | 0.55 ± 0.04ᵇ | 0.41 ± 0.10ᵃᵇ | 0.44 ± 0.15ᵃᵇ | 1.07 ± 0.16ᶜ | 0.07 | 0.000 |
| VFA | 88.30 ± 11.75ᵃ | 124.92 ± 6.05ᵇ | 136.37 ± 4.47ᶜ | 139.02 ± 4.51ᶜᵈ | 146.38 ± 4.85ᵈ | 4.37 | 0.000 |
| IVDMD | 69.65 ± 0.24ᵃ | 72.90 ± 0.55ᵇ | 74.53 ± 0.25ᶜ | 77.60 ± 0.08ᵈ | 78.77 ± 0.65ᵉ | 0.88 | 0.000 |
| IVOMD | 68.58 ± 0.22ᵃ | 72.12 ± 0.57ᵇ | 74.07 ± 0.25ᶜ | 77.28 ± 0.15ᵈ | 78.12 ± 0.70ᵉ | 0.93 | 0.000 |
| IVCPD | 81.96 ± 0.15ᵃ | 83.32 ± 0.24ᵇ | 84.54 ± 0.03ᶜ | 85.29 ± 0.16ᵈ | 88.03 ± 0.62ᵉ | 0.54 | 0.000 |
| IVGED | 59.25 ± 0.33ᵃ | 65.91 ± 0.69ᵇ | 68.68 ± 0.32ᶜ | 69.52 ± 0.11ᶜ | 74.40 ± 0.90ᵈ | 1.33 | 0.000 |
Different superscripts in the same row indicate significant differences (p < 0.05), SEM = Standard error of the means, GC = Green concentrate, GCC1 = GC fortified with 2.5%
Rumen performance parameters were markedly influenced by dietary treatment. Increasing levels of
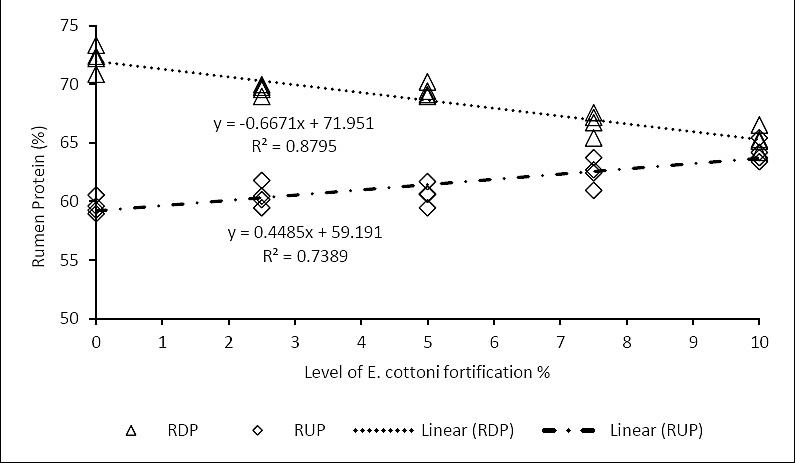
Figure 2. Correlation between
Furthermore, enrichment mitigated methane accumulation, as evidenced by significant reductions in methane production (p = 0.01) and total gas output (p < 0.00) (Table 3). There was a strong correlation (R² = 0.87) between the level of
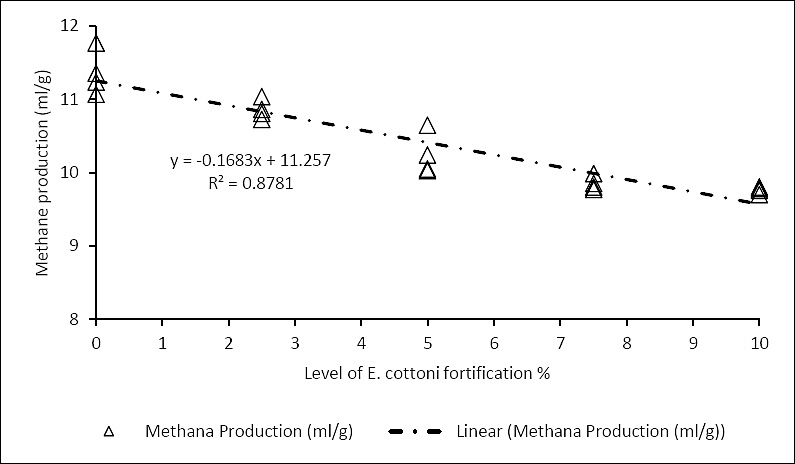
Figure 3. Correlation between
Rumen protein utilization were also significantly altered. Enrichment with
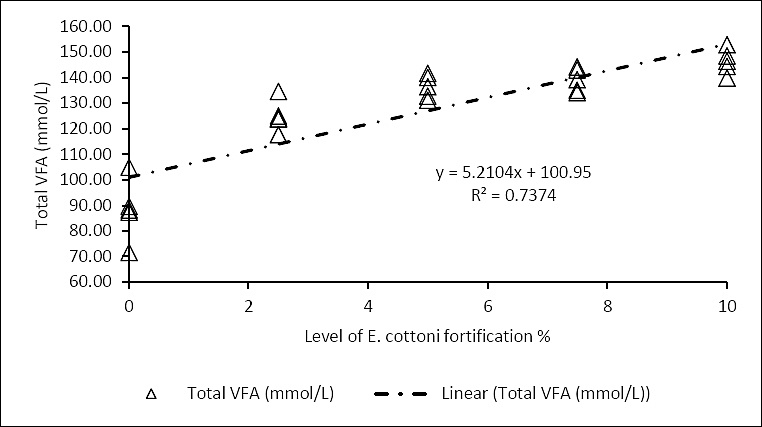
Figure 4. Correlation between
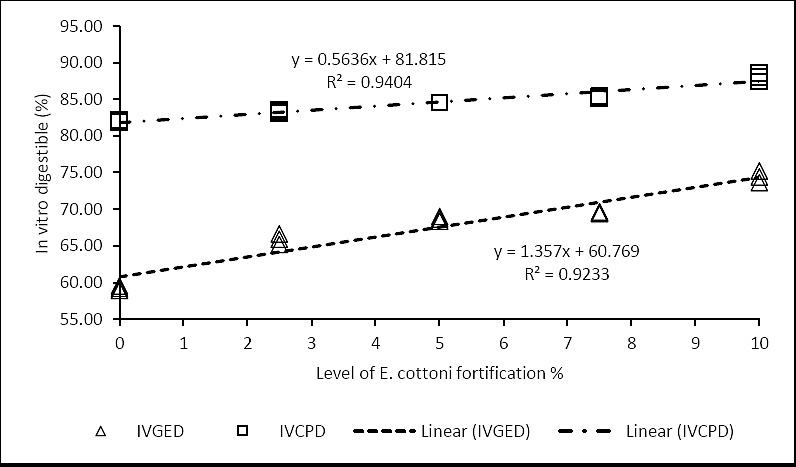
Figure 5. Correlation between
Performance, economic benefit, and milk quality of HF dairy cows fed GC fortified with E. cottonii
Based on Table 4, GC enriched with 10%
Table 4. Performance, milk quality, and economic benefits of Holstein–Friesian dairy cows fed green concentrate fortified with
| Parameters | CON | GC | GCC4 | SEM | p-value |
|---|---|---|---|---|---|
| Production performance | |||||
| Milk yield (kg/day) | 8.67 ± 1.03ᵃ | 9.36 ± 0.49ᵃᵇ | 10.05 ± 0.24ᵇ | 0.20 | 0.011 |
| DMI (kg/day) | 10.63 ± 0.16 | 10.81 ± 0.38 | 10.81 ± 0.32 | 0.07 | 0.496 |
| FCE | 0.81 ± 0.09ᵃ | 0.86 ± 0.03ᵃᵇ | 0.92 ± 0.04ᵇ | 0.01 | 0.022 |
| BCS | 2.50 ± 0.15 | 2.45 ± 0.24 | 2.65 ± 0.12 | 0.04 | 0.177 |
| Economic benefit | |||||
| Feed input (USD/kg DM) | |||||
| Concentrate | 0.45 | 0.30 | 0.33 | – | – |
| Forage | 0.15 | 0.15 | 0.15 | – | – |
| Daily feed cost (USD/day/head) | 2.81 ± 0.02ᶜ | 2.29 ± 0.06ᵃ | 2.43 ± 0.05ᵇ | 0.22 | 0.000 |
| Income from daily production (USD/day/head) | 9.97 ± 1.33ᵃ | 10.76 ± 0.64ᵃᵇ | 11.56 ± 0.31ᵇ | 0.27 | 0.044 |
| Daily profit (USD/day/head) | 7.16 ± 1.31ᵃ | 8.74 ± 0.61ᵇ | 9.13 ± 0.33ᵇ | 0.30 | 0.011 |
| Milk quality | |||||
| Fat (%) | 3.85 ± 0.36ᵃ | 4.03 ± 0.29ᵃ | 5.90 ± 1.20ᵇ | 0.27 | 0.000 |
| Protein (%) | 3.12 ± 0.03ᵃ | 3.13 ± 0.05ᵃ | 3.34 ± 0.08ᵇ | 0.02 | 0.000 |
| Lactose (%) | 4.88 ± 0.22 | 4.82 ± 0.20 | 4.90 ± 0.18 | 0.04 | 0.432 |
| SNF (%) | 8.75 ± 0.28 | 8.78 ± 0.38 | 8.96 ± 0.32 | 0.07 | 0.115 |
| Density (kg/m³) | 1029.91 ± 1.07 | 1029.88 ± 1.32 | 1030.12 ± 0.79 | 0.33 | 0.466 |
| Whey (g/1000 g) | 753.67 ± 16.17ᵇ | 708.37 ± 14.84ᵃ | 710.93 ± 26.19ᵃ | 6.66 | 0.002 |
| Curd (g/1000 g) | 246.32 ± 16.17ᵃ | 291.63 ± 14.84ᵇ | 289.06 ± 26.19ᵇ | 6.66 | 0.002 |
Different superscripts in the same row indicate significant differences (p < 0.05), SEM = Standard error of the means, HF = Holstein Friesian, CON = Commercial concentrate, GC = Green concentrate, GCC4 = GC fortified with 10%
DISCUSSION
Chemical composition of diet
Green concentrates, both enriched and unenriched with
The decline in fat content may also be explained by the hydrophilic nature of seaweed meal, which preferentially binds water rather than fat. Typically, fat molecules are associated with the positive poles of proteins; however, the addition of seaweed meal shifts protein binding affinity toward water, thereby reducing its capacity to bind fat [23]. Conversely, CFi content increased with higher enrichment levels, reflecting the substantial fiber fraction of
Overall, fortification up to 10%
Rumen protein characteristics
The RDP values exhibited a clear decreasing trend with increasing levels of encapsulated
The underlying mechanism is attributed to the presence of kappa-carrageenan from
Without RPA, excess RDP can lead to elevated ammonia (NH3) concentrations, which are absorbed into the bloodstream, converted to urea in the liver, and excreted in urine. Subsequent hydrolysis of urinary urea releases NH3 into the environment, contributing to nitrogen pollution [25]. Thus, the inclusion of kappa-carrageenan from
Beyond its role in protein protection, kappa-carrageenan possesses unique functional properties. Its gel-forming ability has been widely utilized in food systems to protect sensitive nutrients from degradation in harsh environments, including the rumen [27, 28]. Furthermore, kappa-carrageenan from
Overall, the observed decrease in RDP and increase in RUP associated with higher fortification levels confirm the effectiveness of
The RUP refers to the fraction of dietary protein that escapes microbial degradation in the rumen and flows directly to the abomasum and small intestine, where it undergoes enzymatic digestion and absorption. Approximately 80% of RUP is absorbed as amino acids in the small intestine, making it a critical source of high-quality amino acids for highly productive ruminants. These amino acids are essential for supporting milk protein synthesis, metabolic functions, and overall lactation performance. Consequently, diets for high-yielding dairy cows must contain an adequate proportion of RUP to meet post-ruminal amino acid requirements [26].
In the present study, RUP values increased consistently with higher inclusion levels of encapsulated
The mechanism underlying this improvement is based on the content of
Overall, the observed increase in RUP with higher enrichment levels demonstrates the practical and nutritional significance of incorporating
Polysaccharides play a critical role in enhancing RUP by forming stable complexes with proteins, either through chemical interactions such as the Maillard reaction or by acting as physical barriers that slow down proteolysis in the rumen [32]. Among these, chitosan has been widely studied for its ability to increase RUP by creating polymer–protein complexes that reduce microbial access, thereby allowing a greater proportion of dietary protein to bypass ruminal degradation and reach the small intestine for enzymatic digestion [33]. This mechanism improves amino acid availability for absorption and utilization in highly productive ruminants and supports nitrogen pollution reduction quantified via RDP changes.
Kappa-carrageenan, a sulfated polysaccharide abundant in
The practical implications of this mechanism are significant. By reducing ruminal protein degradation and increasing bypass protein availability,
Overall, the data demonstrate that encapsulated
Gas production characteristics
This study demonstrates that enrichment with
Rich in bioactive compounds such as polysaccharides and antioxidants,
These effects collectively enhance short-chain fatty acid synthesis and metabolizable energy availability, ultimately optimizing dairy cow productivity. This innovation has the potential to substantially reduce both ammonia and methane emissions, two key contributors to global warming. Its development represents a strategic effort to advance sustainable and environmentally responsible solutions. Moreover, this approach aligns directly with the aims of the 13th Sustainable Development Goals, which emphasizes the urgent need for climate action.
Rumen performance characteristics
The analysis revealed that enrichment with
The observed increase in VFA concentrations can be attributed to the polysaccharide content of
VFAs are generally classified into two categories: short-chain fatty acids, which represent a major energy source and can supply up to 80% of an animal’s daily energy requirements, including acetate, propionate, and butyrate; and branched-chain VFAs, such as valeric, isobutyric, and isovaleric acids. Branched-chain VFAs play a critical role in stimulating microbial protein synthesis and supporting the proliferation of cellulolytic bacteria, thereby enhancing fiber digestion and overall rumen function [38].
In vitro digestibility of nutrients
As shown in Table 3, increasing enrichment levels corresponded with improved
The mechanism of action is comparable to that of chitosan, offering notable nutritional benefits such as reduced ammonia nitrogen production and enhanced protein bypass to the lower gut [39]. Digestibility serves as a critical indicator of nutrient availability and FCE in ruminants; improved digestibility directly enhances nutrient intake, making it a key determinant of feed effectiveness [5].
Milk yield and FCE
Cows receiving GC and GCC4 produced significantly higher milk yields and exhibited improved FCE compared to those on the control diet (CON). The highest milk yield was recorded in cows fed GCC4, reaching 10.05 kg/day (Table 4). In addition to the benefits of GC, enrichment with
This improvement is attributed to the ability of
Feed quality remains the primary determinant of milk yield in HF dairy cows. As shown in Table 3, GCC4 with 10%
Economic performance and feed affordability
Feed cost represents a major challenge for dairy farmers, as it accounts for approximately 70–80% of total production expenses in dairy farming. Key economic indicators such as feed efficiency, feed cost, milk yield, and milk price significantly influence farm profitability [43].
The inclusion of GC and GCC4 in the diet demonstrates strong potential for adoption by farmers, as these feeding strategies yield higher daily profits compared to the conventional diet (CON). Notably, GCC4 achieved the highest daily profit despite its slightly higher feed cost relative to GC, owing to its superior milk yield performance.
The approach employed in this study emphasizes the utilization of locally available feed resources with optimized nutrient composition and high digestibility through fortification with
Compared with other RPA applications such as chitosan, the use of
Milk quality
The primary components determining milk quality are fat and protein content, both of which are strongly influenced by feed quality. In this study, milk quality complied with the Indonesian National Standard for dairy products. Notably, cows fed GCC4 exhibited higher fat and protein concentrations in milk, indicating improved nutrient absorption and utilization for milk component synthesis [44].
Beyond fat and protein, the quality of whey and curd was also evaluated in Enrekang, where curd yield is of particular economic importance to farmers, as milk is traditionally processed into dangke (a local soft cheese) for sale. GCC4 produced the highest curd yield, attributable to its elevated solids content, including fat, protein, lactose, and SNF.
Milk protein and fat levels are critical predictors of curd yield, as specific thresholds are required to achieve desirable texture and consistency during curd formation. Conversely, whey and curd yields exhibit an inverse relationship; an increase in whey volume typically corresponds to reduced curd production, signaling lower milk solids content [20].
CONCLUSION
The present study demonstrates that enrichment of GC with
From a practical perspective, the use of
A major strength of this study lies in its integrated design combining
However, certain limitations should be acknowledged. The study was conducted under specific environmental and management conditions, which may influence the generalizability of the findings. The sample size, although adequate for detecting treatment effects, remains relatively limited for broader population-level inference. In addition, long-term effects of
Future research should focus on long-term feeding trials to evaluate the sustainability of responses and potential impacts on animal health and reproduction. Further investigation into the molecular and microbiological mechanisms underlying rumen modulation by
In conclusion, enrichment of GC with
DATA AVAILABILITY
The data generated during the study are included in the manuscript.
AUTHORS’ CONTRIBUTIONS
RFU, AA, and ZR: Conceived, designed, and coordinated the study and drafted the manuscript. RFU and ZR: Conducted field sampling, data collection, laboratory work, and data entry. RFU, AA, MMR, JM, and KU: Performed statistical analysis. RFU, ZR, JM, and MMR: Interpreted statistical results. RFU and ZR: Conducted field experiments and tabulated data. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the financial support provided by the Ministry of Higher Education, Science, and Technology of the Republic of Indonesia through the Fundamental-Regular Research Program under Decree No. 0419/C3/DT.05.00/2025 and Agreement/Contract No. 069/C3/DT.05.00/PL/2025. The authors also extend their sincere appreciation to the Dean of the Faculty of Animal Science, Hasanuddin University, and the Department of Animal Husbandry and Fisheries of Enrekang Regency for their valuable assistance and collaboration in facilitating the implementation of this research. We would like to extend our sincere gratitude to Daur Algae for generously providing access to
REFERENCES
- Utamy RF, Ako A, Hasbi H, Ramadan Z, Hakim ARR, Sukri SA. Performance, physiological status, and heat tolerance of Holstein Friesian dairy cows at different lactation phases. Adv. Anim. Vet. Sci 2024;12(10):2034-2042. [Google Scholar]
- Hernández-Castellano LE, Nally JE, Lindahl J, Wanapat M, Alhidary IA, Fangueiro D. Dairy science and health in the tropics: Challenges and opportunities for the next decades. Trop. Anim. Health Prod 2019;51(5):1009-1017. [Google Scholar]
- Sinclair KD, Garnsworthy PC, Mann GE, Sinclair LA. Reducing dietary protein in dairy cow diets: Implications for nitrogen utilization, milk production, welfare and fertility. Animals 2018;8(2):262-274. [Google Scholar]
- Silvi S, Ako A, Utamy RF, Maruddin F, Gustiana S, Umpuch K. Production performance and milk quality of Holstein Friesian dairy cow with the utilization of eggshell meal extract as a source of minerals. Int. J. Agric. Biosci 2024;13(4):626-631. [Google Scholar]
- Utamy RF, Ako A, Ramadan Z, Ishii Y, Rahman MM, Umpuch K. Legume-based green concentrate alleviates negative energy balance and enhances performance, metabolic health, and profitability in postpartum Holstein Friesian cows. Vet. World 2025;18(8):2414-2426. [Google Scholar]
- Utamy RF, Ako A, Ramadan Z, Dagong MIA, Sonjaya H, Maruddin F. Evaluating green concentrate on physiological and metabolic blood profile of Friesian Holstein cows in negative energy balance. Adv. Anim. Vet. Sci 2025;13(5):1113-1123. [Google Scholar]
- Ako A, Utamy RF, Baba S, Hastang, Rahman AA. The effect of leaf meal supplement on milk yield and quality of Friesian Holstein dairy cow. Livest. Res. Rural Dev 2023;35(3). [Google Scholar]
- Ramadan Z, Utamy RF, Hasbi H, Ako A, Maruddin F, Niode V. Economic feasibility and growth performance of Holstein Friesian calves fed whey-dangke fortified green calf starter. Am. J. Anim. Vet. Sci 2025;20(2):159-170. [Google Scholar]
- Situ W, Xiang T, Yan L. Chitosan-based particles for protection of proteins during storage and oral administration. Int. J. Biol. Macromol 2018;117:308-314. [Google Scholar]
- Xie Y, Ding J, Li Y, Wei P, Liu S, Yang R. The formation of protein–chitosan complexes: Their interaction, applications, and challenges. Foods 2024;13:3572. [Google Scholar]
- Harahap RP, Sholikin MM, Sadarman. Chitosan oligosaccharides as dietary antioxidants in nutrition of broiler chickens: A review. Online J. Anim. Feed Res 2024;14(2):107-115. [Google Scholar]
- Ghahfarokhi MG, Barzegar M, Sahari MA, Gavlighi HA, Gardini F. Chitosan-cinnamon essential oil nano-formulation: Application as a novel additive for controlled release and shelf life extension of beef patties. Int. J. Biol. Macromol 2017;102:19-28. [Google Scholar]
- Anggraini J, Lo D. Health impact of carrageenan and its application in food industry: A review. IOP Conf. Ser. Earth Environ. Sci 2023;1169:012098. [Google Scholar]
- Natsir A, Pasia R, Hidayat R, Ginting N, Ismartoyo I, Syahrir S. Enhancing rumen efficiency and reducing methane emissions with South Sulawesi seaweed as feed additives. Int J Agric Biosci 2025;14(4):529-538. [Google Scholar]
- Latimer GW. Official methods of analysis of AOAC International. Gaithersburg, MD: AOAC International; 2019. [Google Scholar]
- Tilley JMA, Terry RAA. two-stage technique for the
in vitro digestion of forage crops. Grass For. Sci 1963;18(2):104-111. [Google Scholar] - Ørskov ER, McDonald I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci 1979;92:499-503. [Google Scholar]
- Haryati RP, Jayanegara A, Laconi EB, Ridla M, Suptijah P. Evaluation of chitin and chitosan from insect as feed additives to mitigate ruminal methane emission. AIP Conf. Proc 2019;2120:040008. [Google Scholar]
- Edmonson AJ, Lean IJ, Weaver LD, Farver T, Webster G. A body condition scoring chart for Holstein dairy cows. J. Dairy Sci 1989;72:68-78. [Google Scholar]
- Syawal S, Utamy RF, Ako A, Hasbi H, Rahman AA, Ramadan Z. Milk quality, curd, and whey percentages of Holstein Friesian cows at different lactation phases. Hasanuddin J. Anim. Sci 2025;7(1):56-64. [Google Scholar]
- Nutrient requirements of dairy cattle. Washington, DC: National Academy Press; 2001. [Google Scholar]
- Kaseng ES, Suhaeb FW. Analysis of the effect of seaweed (
Eucheuma cottonii ) as substitution material to tapioca flour on the quality of snakehead fish (Channa striata ) nuggets. J. Phys. Conf. Ser 2021;1752:012054. [Google Scholar] - Fiqri F, Junianto J, Maulina I, Rostini I. Effect of seaweed (
Eucheuma cottonii ) flour addition on preference level of choux pastry. J. Fish Health 2025;5(3):318-330. [Google Scholar] - Permana IG, Rosmalia A, Anggarini FY, Despal D, Toharmat T, Evvyernie D. Optimizing tropical dairy goat diets: Balancing rumen degradable protein, non-fiber carbohydrates, and sulfur requirements. Anim. Biosci 2025;38(3):475-488. [Google Scholar]
- Savari M, Khorvash M, Amanlou H, Ghorbani GR, Ghasemi E, Mirzaei M. Effects of rumen-degradable protein: Rumen-undegradable protein ratio and corn processing on production performance, nitrogen efficiency, and feeding behavior of Holstein dairy cows. J. Dairy Sci 2017;101:1111-1122. [Google Scholar]
- Putri EM, Zainn M, Warly L, Hermon H. Effects of rumen-degradable-to-undegradable protein ratio in ruminant diet on
in vitro digestibility, rumen fermentation, and microbial protein synthesis. Vet. World 2021;14(3):640-648. [Google Scholar] - Malav OP. Application of carrageenan in food and feed systems. Food Hydrocoll 2015;43:265-271. [Google Scholar]
- Singh A. Encapsulation of nutrients using carrageenan: A review. Int. J. Biol. Macromol 2015;75:451-458. [Google Scholar]
- Xiao Y, He X, Ma J, Dua C, Gan S, Yin F. κ-carrageenan oligosaccharides alleviate MDP-induced rumen epithelial cell inflammatory damage by inhibiting activation of NOD2/NF-κB pathway. Front. Vet. Sci 2025;12:1626423. [Google Scholar]
- Cheong KL, Zhang Y, Li Z, Li T, Ou Y, Shen J. Role of polysaccharides from marine seaweed as feed additives for methane mitigation in ruminants: A critical review. Polymers 2023;15:3153. [Google Scholar]
- Garba AM, Firincioğlu SY. Role of encapsulation nutrients for improvement of ruminant performance and ruminant-derived products. Eurasian J. Agric. Res 2023;7(2):109-126. [Google Scholar]
- Loregian KE, Silva MJ, Dourado SB, Guimarães J, Amâncio BR, Magnani E. Enhancing rumen-undegradable protein via processing techniques in a dual-flow continuous culture system. Fermentation 2025;11:94. [Google Scholar]
- Chang J, Zeng Y, Tao H, Hu D, Zhao W, Wang S. Effects of polysaccharides on protein
in vitro digestion and rheological properties under simulated elderly gastric conditions. Food Chem 2025;492:145662. [Google Scholar] - Herdiana Y. Polymeric rumen-stable delivery systems for delivering nutritions. Open Vet. J 2015;15(2):565-593. [Google Scholar]
- Króliczewska B, Pecka-Kiełb E, Bujok J. Strategies used to reduce methane emissions from ruminants: Controversies and issues. Agriculture 2023;13:602. [Google Scholar]
- Widiawati Y, Widodo S, Adegbeye MJ, Saputra F, Alvarado Ramírez ER, Anggraeny YN. Potential of seaweed (
Eucheuma cottonii ) supplementation to reduce methane production, improve fermentation, and modulate microbial composition duringin vitro rumen fermentation. Front. Vet. Sci 2025;12:1-18. [Google Scholar] - Widiawati Y, Hikmawan D. Enteric methane mitigation by using seaweed
Eucheuma cottonii . IOP Conf. Ser. Earth Environ. Sci 2021;788:012152. [Google Scholar] - Mazinani M, Naserian AA, Rude BJ, Tahmasbi AM, Valizadeh R. Effects of feeding rumen-protected amino acids on performance of feedlot calves. J. Adv. Vet. Anim. Res 2020;7(2):229-233. [Google Scholar]
- Shah AM, Qazi IH, Matra M, Wanapat M. Role of chitin and chitosan in ruminant diets and their impact on digestibility, microbiota, and performance. Fermentation 2022;8(10):2-19. [Google Scholar]
- Álvarez C, Nielsen NI, Weisbjerg MR, Volden H, Eknæs M, Prestløkken E. High-digestible silages allow low concentrate supply without affecting milk production or methane emissions. J. Dairy Sci 2022;105(4):3633-3647. [Google Scholar]
- Chiang YW, Wang TH, Lee WC. Chitosan coating for protection of amino acids entrapped within hydrogenated fat. Food Hydrocoll 2009;23(3):1057-1061. [Google Scholar]
- Casper DP, Pretz JP, Ramsier C. Lactational performance of early-lactation dairy cows fed forages produced by two different crop production programs. Animals 2025;15(13):2-19. [Google Scholar]
- Atzori AS, Valsecchi C, Manca E, Masoero F, Cannas A, Gallo A. Assessment of feed and economic efficiency of dairy farms based on multivariate aggregation of partial indicators measured on field. J. Dairy Sci 2021;104(12):12679-12692. [Google Scholar]
- Eikanger KS, Eknæs M, Sommerseth JK, Karlengen IJ, Schei I, Silberberg M. Feed intake, milk production, and nutrient use efficiency with early lactating Norwegian Red dairy cows fed concentrates with varying levels of local ingredients. Livest. Sci 2025:1-9. [Google Scholar]