


ABSTRACT
Background and Aim:
Materials and Methods: A total of 689 swab samples were collected from two slaughterhouses and two retail markets in Khon Kaen Province, Thailand, across three seasons (summer, rainy, and winter) from March 2023 to February 2024. Isolation and identification of
Results: The overall prevalence of
Conclusion: This study provides a comprehensive, year-round comparative assessment of
Keywords: antimicrobial resistance, chicken, food safety, genetic relatedness, prevalence, poultry supply chain, seasonal variation,
INTRODUCTION
The widespread use of antimicrobials for therapy, prophylaxis, and growth promotion in animal production has been a key driver in the selection and dissemination of antimicrobial resistance (AMR) [6, 7]. The emergence of multidrug-resistant (MDR)
In Thailand, national surveillance data consistently identify non-typhoidal
In Southeast Asia, including countries such as Thailand, large-scale poultry production and the widespread use of antimicrobial agents have made the region a hotspot for AMR development. However, most existing monitoring studies have notable limitations. Firstly, many studies predominantly focus on isolated segments of the production chain, either farms/slaughterhouses or retail markets, thus lacking continuous, systematic comparisons across the entire post-harvest “slaughterhouse-to-market” distribution chain. This hinders the elucidation of dynamic changes in
Despite extensive documentation of
Furthermore, while phenotypic resistance profiles are widely reported, there is limited incorporation of molecular epidemiological tools to assess the genetic relatedness of circulating
Therefore, the present study was designed to provide a comprehensive, integrated assessment of
By integrating epidemiological, phenotypic, and molecular analyses across both spatial (supply chain stages) and temporal (seasonal) dimensions, this study seeks to elucidate critical risk factors driving
MATERIALS AND METHODS
Ethical approval
The study protocol was reviewed and approved by the Institutional Animal Care and Use Committee of Khon Kaen University, Thailand (approval no. IACUC-KKU-111/67). All procedures involving animals were conducted in strict accordance with national guidelines for animal research and welfare, and in compliance with internationally accepted standards, including the ARRIVE 2.0 guidelines and the principles outlined in the Guide for the Care and Use of Laboratory Animals.
This study was based on non-invasive sampling procedures conducted under routine surveillance conditions. Cloacal swab samples were collected by trained veterinarians using standardized handling techniques to minimize stress, discomfort, and potential injury to the birds. No experimental infection, invasive intervention, or euthanasia was performed as part of this study. All sampling procedures adhered to the principles of replacement, reduction, and refinement (3Rs), ensuring that the number of animals involved was limited to the minimum required to achieve scientific validity while maintaining high standards of animal welfare.
The chickens included in this study originated from commercial farms operating under the Thai Agricultural Standard TAS 6901-2017 and were handled within standard production and processing environments. Permission to access animals and collect samples at slaughterhouses and retail markets was obtained from facility operators and relevant authorities prior to sampling. All sampling activities were carried out in a manner that did not interfere with routine farm or slaughterhouse operations.
Biosafety and biosecurity measures were strictly implemented throughout the study. All laboratory procedures involving
Overall, the study design ensured full compliance with ethical standards for animal use in research while maintaining scientific rigor and data reliability.
Study period and location
This study was a cross-sectional seasonal surveillance study conducted along the poultry supply chain in Khon Kaen Province, Thailand. Over one year (March 2023 to February 2024), we simultaneously monitored and compared
Description of slaughterhouses and markets
The chickens sampled in this study originated from standardized commercial farms operating under the Thai Agricultural Standard TAS 6901-2017, which mandates veterinary oversight and restricts the use of certain critically important antimicrobials, and which had been approved by the Department of Livestock Development. All broilers were of a mixed breed, aged 35–40 days, and sourced from a single integrated production company. This common origin ensures a consistent baseline of farm management and biosecurity. Birds were transported to the slaughterhouses in dedicated vehicles over a short travel duration (1–2 h). The two participating slaughterhouses were small scale, manually operated facilities processing approximately 500–800 birds per day, utilizing municipal water and implementing basic daily cleaning routines. Following processing, carcasses were distributed to the two traditional wet markets included in the study, where they were displayed and sold at ambient temperature on open stalls, with no dedicated cold chain facilities.
Sampling strategy and seasonal classification
A total of 689 swabs (including cloacal swab and carcass swab) were collected during different seasons from slaughterhouses (n = 2) and markets (n = 2) in Khon Kaen Province, Northeast Thailand. Three seasons were defined as summer (March–May, 32°C–38°C, humidity rate 66%–82%), rainy (June–October, 27°C–30°C, humidity rate 82%–88%), and winter (November–February, 20°C–28°C, humidity rate 50%–75%). The sampling period spanned one year (March 2023 to February 2024), covering a temperature–humidity index range of 65–80, representing 16 discrete intervals.
Sample size determination
A systematic, repeated sampling strategy was employed: the same four sites were sampled once per quarter across all three seasons. Based on a reported
Table 1. Number of samples collected from slaughterhouses and markets across different seasons.
| Season | Sampling site | Number of samples | Sample type |
|---|---|---|---|
| Summer | Slaughterhouse 1 | 57 | Cloacal swab |
| Summer | Slaughterhouse 2 | 44 | Cloacal swab |
| Summer | Market 1 | 34 | Carcass swab |
| Summer | Market 2 | 33 | Carcass swab |
| Rainy | Slaughterhouse 1 | 78 | Cloacal swab |
| Rainy | Slaughterhouse 2 | 88 | Cloacal swab |
| Rainy | Market 1 | 71 | Carcass swab |
| Rainy | Market 2 | 55 | Carcass swab |
| Winter | Slaughterhouse 1 | 100 | Cloacal swab |
| Winter | Slaughterhouse 2 | 66 | Cloacal swab |
| Winter | Market 1 | 49 | Carcass swab |
| Winter | Market 2 | 77 | Carcass swab |
| Total | 689 |
Isolation and identification of Salmonella
Isolation and identification of
Serotyping of Salmonella isolates
Serological identification was performed using diagnostic polyvalent and monovalent
Antimicrobial susceptibility testing
The panel included agents with reported usage in Thai poultry (e.g., ampicillin, tetracycline, streptomycin) alongside representatives of the WHO highest-priority critically important antimicrobials (HPCIA) (e.g., fluoroquinolones, third-generation cephalosporins, carbapenems). Colistin was excluded based on its restricted use in Thai food animals following the 2017 national ban on antimicrobial growth promoters.
Antimicrobial susceptibility testing was performed against 14 antibiotics using the disk diffusion method on Mueller–Hinton agar (Difco), in accordance with Clinical and Laboratory Standards Institute (CLSI) guidelines [20]. The antimicrobial agents (Oxoid, Liofilchem, UK) tested included streptomycin (S, 10 μg), norfloxacin (NOR, 10 μg), tetracycline (TE, 30 μg), gentamicin (GM, 10 μg), sulfamethoxazole/trimethoprim (SXT, 23.75 μg), imipenem (IPM, 10 μg), cefotaxime (CRO, 30 μg), chloramphenicol (C, 30 μg), ciprofloxacin (CIP, 5 μg), nalidixic acid (NA, 30 μg), ampicillin (AM, 10 μg), ceftazidime (CAZ, 30 μg), ceftriaxone (CTX, 30 μg), and amoxicillin/clavulanic acid (AMC, 20/10 μg). Escherichia coli ATCC 25922 (ATCC, Manassas, VA, USA) was used for quality control. Results were accepted only when the zone diameters for the quality control strain fell within CLSI-specified reference ranges. Multidrug resistance was defined as resistance to agents from three or more distinct antibiotic classes [21].
PFGE and cluster analysis
To investigate the genetic relatedness of the dominant
PFGE profiles were analyzed using BioNumerics software (version 4.5, Bio-Rad). Dendrograms were constructed using the unweighted pair group method with arithmetic mean, with the Dice similarity coefficient, optimization set to 1%, and tolerance set to 1%. Isolates with 100% similarity were considered to belong to the same PFGE type.
Statistical analysis
Data were processed using Excel, SPSS 20.0 (IBM Corp., Armonk, NY, USA), and GraphPad Prism 10.1.3. Categorical data were analyzed using the chi-square test, with statistical significance set at p < 0.05 and high statistical significance at p < 0.01.
The prevalence of
RESULTS
Salmonella prevalence in slaughterhouses and markets with seasonal changes
This study found an overall
Table 2. Seasonal distribution of slaughterhouses and markets from chicken.
| Source | Numbers | Positive (%) | Summer (%) | Rainy (%) | Winter (%) | χ² | P1 value |
|---|---|---|---|---|---|---|---|
| Slaughterhouse 1 | 235 | 54 (22.97) | 7 (2.99) | 29 (12.34) | 18 (7.66) | 15.15 | 0.0005 |
| Slaughterhouse 2 | 181 | 33 (18.23) | 6 (3.31) | 20 (11.05) | 7 (3.87) | 10.12 | 0.0006 |
| Market 1 | 120 | 73 (60.83) | 11 (9.16) | 25 (20.83) | 37 (30.83) | 15.70 | 0.0004 |
| Market 2 | 153 | 63 (41.18) | 15 (9.80) | 27 (17.65) | 21 (13.74) | 3.43 | 0.18 (ns) |
| Total | 689 | 223 (32.37) | 39 (17.49) | 101 (45.29) | 83 (37.22) | – | – |
| χ² (spatial/seasonal) | – | 25.48 | 4.90 | 9.25 | 28.27 | – | – |
| P2 value | – | <0.0001 | 0.175 (ns) | 0.026 | <0.0001 | – | – |
P1 values represent chi-square goodness-of-fit tests for seasonal distribution uniformity of positive isolates within each sampling site. P2 values indicate chi-square goodness-of-fit tests for spatial distribution uniformity of positive isolates across sampling sites within each seasonal category.
To quantify the contamination risk across seasons, this study calculated the RR for the rainy and winter seasons, using summer as the reference. The analysis was stratified by sample type, revealing distinct seasonal risk patterns depending on the sample source. For anal swabs (reflecting animal carriage), the risk of
Table 3.
| Sample category | Season | Total | Positive | Prevalence (%) | RR | 95% CI | p-value |
|---|---|---|---|---|---|---|---|
| Summer | 101 | 13 | 12.87 | 1(ref) | – | – | |
| Anal swab | Rainy | 166 | 49 | 29.52 | 2.29 | 1.32–4.00 | 0.003 |
| Winter | 166 | 25 | 15.06 | 1.17 | 0.63–2.17 | 0.617 | |
| Summer | 67 | 26 | 38.81 | 1(ref) | – | – | |
| Carcass swab | Rainy | 126 | 52 | 41.27 | 1.06 | 0.75–1.51 | 0.732 |
| Winter | 126 | 58 | 46.03 | 1.19 | 0.86–1.65 | 0.299 |
RR uses summer as the reference group; 95% CI denotes 95% confidence interval; P-values are calculated based on chi-square test or Fisher’s exact test.
To assess the independent effects of each risk factor, this study further constructed a multivariate logistic regression model. After adjusting for the confounding effects of season and sampling location, the analysis revealed that retail markets (compared to slaughterhouses) and the rainy season and winter (compared to summer) each represented independent risk factors for
Table 4. Multivariable logistic regression analysis of independent risk factors for
| Factor | Category | Adjusted odds ratio | 95% Confidence interval | p-value |
|---|---|---|---|---|
| Location type | Slaughterhouse (Ref) | 1 | – | – |
| Market | 4.82 | 3.42 – 6.79 | <0.001 | |
| Summer (Ref) | 1 | – | – | |
| Season | Rainy | 3.81 | 2.40 – 6.04 | <0.001 |
| Winter | 2.80 | 1.73 – 4.53 | <0.001 |
Model fit statistics: Hosmer-Lemeshow goodness-of-fit test, χ² = 5.32, p = 0.62; Nagelkerke R² = 0.28. The overall model was statistically significant (
Serotype distribution from slaughterhouses and markets
Serotyping of 223
Table 5. Serotype distribution of different isolates from slaughterhouses and markets.
| Serotype | Slaughterhouse 1 | Slaughterhouse 2 | Market 1 | Market 2 | Season | Total | |
|---|---|---|---|---|---|---|---|
| 17 (26.98) | 14 (22.58) | 2 (3.85) | 0 | Rainy | 33 (14.79) | ||
| 13 (20.63) | 17 (27.42) | 2 (3.85) | 2 (4.76) | Winter | 34 (15.25) | ||
| 1 (0.16) | 0 | 8 (15.38) | 2 (4.76) | Rainy | 11 (0.49) | ||
| B | 2 (0.32) | 0 | 1 (1.92) | 0 | Winter | 3 (0.13) | |
| 0 | 0 | 2 (3.85) | 1 (2.38) | Winter | 3 (0.13) | ||
| 0 | 0 | 1 (1.92) | 0 | Rainy | 1 (0.04) | ||
| 0 | 0 | 2 (3.85) | 2 (4.76) | Rainy | 4 (0.18) | ||
| 0 | 2 (3.23) | 3 (5.77) | 3 (7.14) | Rainy | 8 (0.36) | ||
| 5 (7.94) | 7 (11.29) | 8 (15.38) | 3 (7.14) | Rainy | 23 (10.31) | ||
| 5 (7.94) | 3 (4.84) | 0 | 0 | Rainy | 8 (0.36) | ||
| 4 (6.35) | 6 (9.68) | 0 | 0 | Rainy | 1 (0.45) | ||
| 1 (1.59) | 0 | 0 | 0 | Rainy | 1 (0.04) | ||
| C | 1 (1.59) | 4 (6.45) | 7 (13.46) | 4 (9.52) | Rainy | 16 (0.72) | |
| 1 (1.59) | 2 (3.23) | 2 (3.85) | 5 (11.9) | Rainy | 1 (0.45) | ||
| 1 (1.59) | 0 | 10 (19.23) | 7 (16.66) | Winter | 18 (0.81) | ||
| 1 (1.59) | 0 | 3 (5.77) | 5 (11.9) | Winter | 9 (0.40) | ||
| 0 | 0 | 0 | 1 (2.38) | Winter | 1 (0.04) | ||
| 1 (1.59) | 1 (1.61) | 3 (5.77) | 0 | Winter | 5 (0.22) | ||
| 0 | 0 | 0 | 1 (2.38) | Rainy | 1 (0.04) | ||
| 0 | 0 | 0 | 1 (2.38) | Winter | 1 (0.04) | ||
| D | 6 (9.52) | 3 (4.84) | 0 | 0 | Winter | 9 (0.40) | |
| E | 0 | 0 | 0 | 1 (2.38) | Rainy | 1 (0.04) | |
| I | 1 (1.59) | 0 | 0 | 0 | Summer | 1 (0.04) | |
| 3 (4.76) | 3 (4.84) | 2 (3.85) | 4 (9.52) | Winter | 1 (0.54) | ||
| Total | 63 | 62 | 52 | 42 | 223 |
Among positive isolates,
Table 6. The seasonal variation of
| Season | Positive | Agona | Non-Agona | Rate (%) | χ² | p-value |
|---|---|---|---|---|---|---|
| Summer | 38 | 2 | 36 | 5.56 | ||
| Rainy | 101 | 9 | 92 | 9.78 | 15.64 | <0.001 |
| Winter | 84 | 23 | 61 | 37.70 |
Chi-square test (df = 2) was performed on the distribution of
Antibiotic resistance profile
The prevalence of AMR (to at least one drug) among
The heat map (Figure 1A) indicates that isolates from all locations exhibited the highest resistance rates to streptomycin (S) (39.7%–50.0%), with particularly pronounced resistance to ampicillin (AM) at Market-2 (57.1%) and Market-1 (35.6%); Slaughterhouse-1 exhibited significantly higher resistance rates to gentamicin (GM, 37.0%) and NA (48.1%) compared to other sites, whereas no resistance was detected to critically important antimicrobials such as CIP, NOR, or IMP. This illustrates severe resistance to first-line drugs, with distinct resistance patterns across locations, suggesting that the emergence and persistence of AMR are closely linked to site-specific environmental factors.
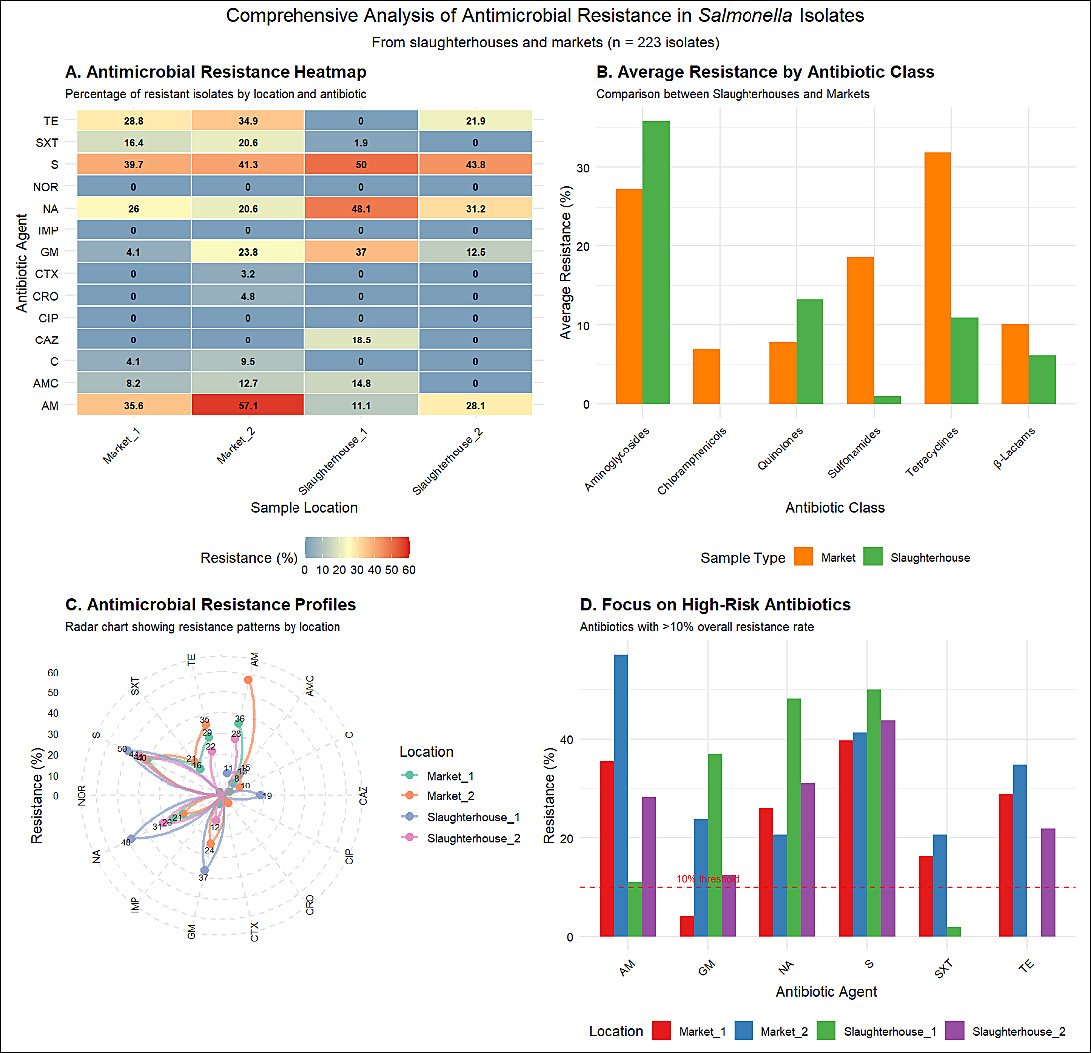
Figure 1. Analysis of antimicrobial resistance in
Comparisons revealed that isolates from market sources exhibited higher average resistance rates, particularly to β-lactams and tetracyclines than those from slaughterhouse sources, while isolates from slaughterhouse sources demonstrated higher average resistance rates to aminoglycosides and quinolones (Figure 1B). This indicates distinct antimicrobial selection pressures along the supply chain from slaughterhouses to markets, potentially reflecting variations in antimicrobial usage practices or contamination sources at different stages.
Radar charts revealed unique AMR patterns across locations (Figure 1C), with Slaughterhouse-1 exhibiting the broadest resistance spectrum (the highest levels of multidrug resistance) and Market-2 showing particularly pronounced resistance to certain antimicrobial classes (e.g., β-lactams). These findings confirm significant location-specific resistance profiles, underscoring the need for targeted prevention and control strategies tailored to different settings rather than generic universal approaches.
Analysis of high-risk drugs with overall resistance rates >10% confirmed streptomycin (S), ampicillin (AM), NA, and tetracycline (TE) as primary threats. It clearly showed that Slaughterhouse-1 is the main location contributing to high resistance rates for gentamicin (GM) and NA, while Market-2 is a hotspot for ampicillin (AM) and tetracycline (TE) resistance (Figure 1D). This analysis identifies high-risk antibiotics requiring priority intervention and their core sources, providing direct evidence for implementing targeted surveillance and control measures.
To further investigate the distribution of MDR
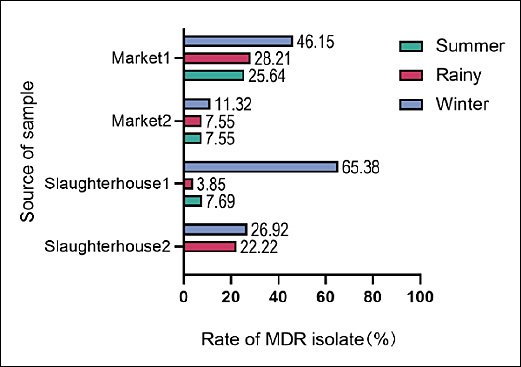
Figure 2. Seasonal and spatial distribution of multidrug-resistant (MDR)
Genetic diversity and AMR patterns of Salmonella isolates
To explore potential clonal relationships and provide genetic context for the observed spatiotemporal and phenotypic patterns, all available isolates of the most prevalent serotype,
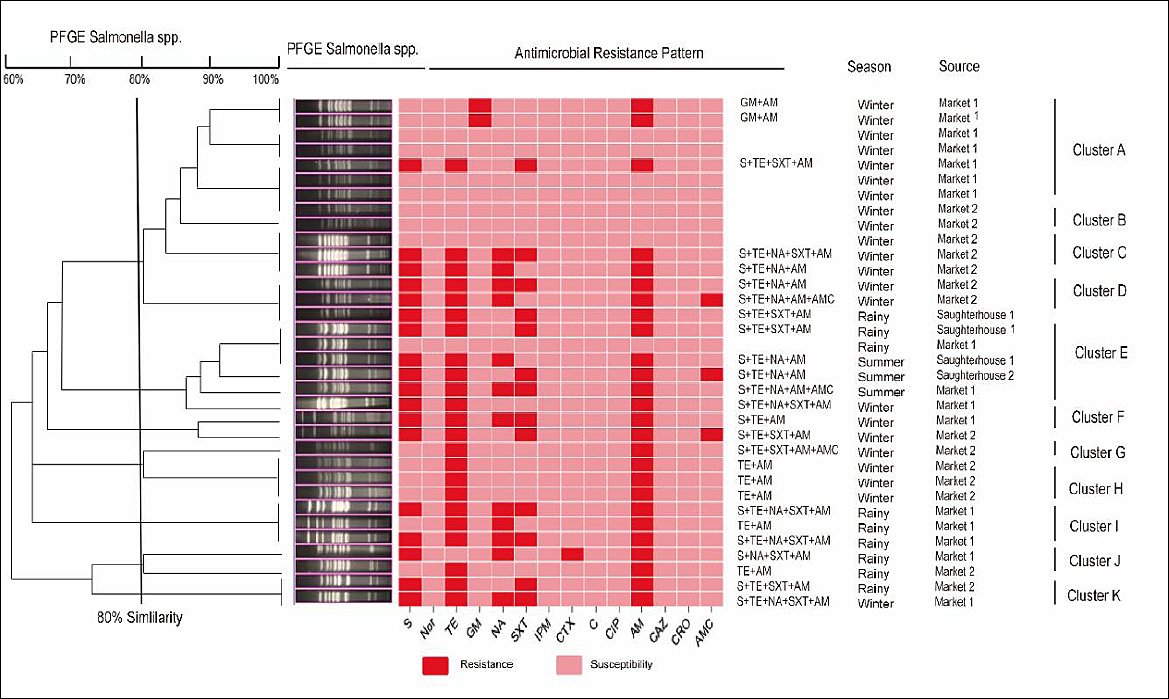
Figure 3. Genetic relatedness of 34
DISCUSSION
Overall prevalence and supply chain contamination dynamics
The present study provides a comprehensive overview of the prevalence, serotype distribution, AMR, and genetic relatedness of
Seasonal variation and environmental drivers
The significant seasonal variation observed, with the highest prevalence during the rainy season (45.29%), parallels findings from tropical regions worldwide [28]. Studies conducted in Bangladesh demonstrated similar seasonal patterns where monsoon conditions facilitated
Seasonal dissociation between prevalence and multidrug resistance
The most novel and operationally relevant finding of this study is the distinct seasonal dissociation between peak
Temporal shift in serotype distribution
Comparison with historical data from the same province reveals a notable shift in serotype predominance over the past two decades. While Angkititrakul
Serotype ecology and persistence mechanisms
The predominance of
AMR patterns and regional context
While direct farm-level antimicrobial usage (AMU) data were not available for correlation with our findings, the resistance patterns observed can be contextualized within the known landscape of AMU in Thai poultry production as reported in the literature. The resistance profiles observed, high rates to streptomycin (43.05%), ampicillin (34.53%), and NA (30.49%), are consistent with national and regional surveillance data from Thailand and neighboring countries [38–40]. The rising resistance gradient from slaughterhouses to markets parallels findings from Vietnam, suggesting additional selective pressure or cross-contamination at the retail level [41].
Critically important antimicrobials and regulatory impact
The absence of resistance to fluoroquinolones and carbapenems is encouraging and likely reflects Thailand’s 2017 colistin ban and subsequent restrictions on highest-priority critically important antimicrobials in food animals [42]. The notably higher ampicillin resistance in markets (57.14%) compared to slaughterhouses (11.11%) may reflect unauthorized farm-level antibiotic use or cross-contamination during market handling, a pattern observed elsewhere in the region [43].
MDR dynamics and transmission pathways
The high MDR prevalence in slaughterhouses (65.38% in Slaughterhouse-1), peaking in winter, highlights slaughterhouses as critical nodes for resistance amplification. This seasonal pattern may reflect prophylactic antibiotic use during cold months or flock consolidation prior to winter slaughter [44, 45]. PFGE clustering of isolates by source and season supports localized clonal dissemination, while shared PFGE types across sites suggest potential cross-contamination along the supply chain [46]. These findings reinforce the need for integrated One Health surveillance that links human salmonellosis cases with poultry supply chain data to better inform public health interventions.
Study limitations and future research directions
Several limitations of the present study should be acknowledged. First and foremost, this study did not collect AMU data from the primary source farms. Consequently, critical information regarding the specific types of antimicrobials used, the distinction between therapeutic and prophylactic applications, and potential seasonal variations in antimicrobial medication practices remains unknown. Without these farm-level AMU records, any robust causal inference between antimicrobial exposure and the observed resistance patterns remains speculative. Second, the absence of comprehensive genotypic characterization of resistance determinants limits our understanding of the underlying resistance mechanisms and their potential for horizontal gene transfer. Third, environmental samples were not collected from sampling sites, restricting our capacity to trace cross-contamination pathways. Fourth, PFGE offers lower discriminatory power than whole-genome sequencing for fine-scale bacterial transmission tracing. Fifth, the cross-sectional design only captures associations at discrete time points but cannot establish temporal causality. Finally, the single-province scope may limit the generalizability of the findings to other regions of Thailand. Despite these limitations, this study provides valuable, much-needed baseline data on the spatiotemporal dynamics of
CONCLUSION
This study provides a comprehensive assessment of
From a practical perspective, these findings provide direct evidence to support targeted, season-specific intervention strategies. Enhanced hygiene, water management, and biosecurity measures should be prioritized during the rainy season to reduce environmental contamination, while winter interventions should focus on antimicrobial stewardship and monitoring of MDR within slaughterhouses to limit the amplification and dissemination of resistant strains. The identification of slaughterhouses as critical nodes for MDR emergence further emphasizes the need for strengthened control measures at this stage of the supply chain.
A major strength of this study lies in its integrated design, combining seasonal surveillance, comparative analysis across supply chain nodes, AMR profiling, and pulsed-field gel electrophoresis-based genetic characteri-zation. This approach enabled the identification of spatial–temporal risk patterns and provided molecular evidence supporting localized clonal dissemination and potential cross-contamination pathways.
Nevertheless, future research should address key limitations by incorporating longitudinal farm-level antimicrobial usage data, environmental sampling, and high-resolution genomic approaches such as whole-genome sequencing to better elucidate transmission dynamics and resistance mechanisms. Expanding surveillance across multiple regions would also improve the generalizability of the findings.
In conclusion,
DATA AVAILABILITY
Representative original gel images (PFGE) and representative photographs (AST) are provided in the Supplementary Materials. The complete set of original image files is available from the corresponding author upon reasonable request.
AUTHORS’ CONTRIBUTIONS
ZZ: Investigation, data curation, formal analysis, writing – original draft, and visualization. FS: Methodology, validation, investigation, and writing – review and editing. CP: Resources, software, supervision, and writing – review and editing. SA: Conceptualization, funding acquisition, project administration, supervision, and writing – review and editing. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors would like to thank the Veterinary Public Health Platform, Faculty of Veterinary Medicine, Khon Kaen University, Thailand, for providing technical support and facilities that greatly facilitated this study. The authors also gratefully acknowledge the KKU Scholarship for ASEAN and GMS Countries’ Personnel of Academic Year 2023 (No. 730/2023) for financial support.
REFERENCES
- EFSA J 2021;19:e06406. [Google Scholar] | [Crossref]
- Balasubramanian R, Im J, Lee J-S, Jeon HJ, Mogeni OD, Kim JH. The global burden and epidemiology of invasive non-typhoidal
Salmonella infections. Hum Vaccin Immunother 2019;15(6):1421-6. [Google Scholar] | [Crossref] - Jechalke S, Schierstaedt J, Becker M, Flemer B, Grosch R, Smalla K.
Salmonella establishment in agricultural soil and colonization of crop plants depend on soil type and plant species. Front Microbiol 2019;10:967. [Google Scholar] | [Crossref] - Koenig A, Cooper TL, Greene CE, Sharma A, Sakamoto K. Clinical salmonellosis in a closed colony of blood donor cats. Comp Med;2017(67):524-8. [Google Scholar] | [Crossref]
- Galán-Relaño Á, Valero Díaz A, Huerta Lorenzo B, Gómez-Gascón L, Mena Rodríguez MÁ, Carrasco Jiménez E.
Salmonella and salmonellosis:an update on public health implications and control strategies. Animals (Basel) 2023;13(23):3666. [Google Scholar] | [Crossref] - Zeng H, De Reu K, Gabriël S, Mattheus W, De Zutter L, Rasschaert G.
Salmonella prevalence and persistence in industrialized poultry slaughterhouses. Poult Sci 2021;100(3):100991. [Google Scholar] | [Crossref] - Ajulo S, Awosile B. Global Antimicrobial Resistance and Use Surveillance System (GLASS 2022):investigating the relationship between antimicrobial resistance and antimicrobial consumption data across the participating countries. PLoS One 2024;19(2):e0297921. [Google Scholar] | [Crossref]
- Kumar H, Chen B-H, Kuca K, Nepovimova E, Kaushal A, Nagraik R. Understanding of colistin usage in food animals and available detection techniques:a review. Animals (Basel) 2020;10(10):1892. [Google Scholar] | [Crossref]
- Antunes P, Mourão J, Campos J, Peixe L. Salmonellosis:the role of poultry meat. Clin Microbiol Infect 2016;22(2):110-21. [Google Scholar] | [Crossref]
- Brown D. Antibiotic resistance breakers:can repurposed drugs fill the antibiotic discovery void?. Nat Rev Drug Discov 2015;14(12):821-32. [Google Scholar] | [Crossref]
- Nhung NT, Phu DH, Carrique-Mas JJ, Padungtod P. A review and meta-analysis of non-typhoidal
Salmonella in Vietnam:challenges to the control and antimicrobial resistance traits of a neglected zoonotic pathogen. One Health 2024;18:100698. [Google Scholar] | [Crossref] - Chotinun S, Rojanasthien S, Unger F, Tadee P. Prevalence and antimicrobial resistance of
Salmonella isolated from carcasses, processing facilities and the environment surrounding small scale poultry. Southeast Asian J Trop Med Public Health 2014;45(6):1571-81. [Google Scholar] | [Crossref] - Prestinaci F, Pezzotti P, Pantosti A. Antimicrobial resistance:a global multifaceted phenomenon. Pathog Glob Health 2015;109(7):309-18. [Google Scholar] | [Crossref]
- Eiamsam-Ang T, Tadee P, Pascoe B, Patchanee P. Genome-based analysis of infrequent
Salmonella serotypes through the Thai pork production chain. Front Microbiol 2022;13:968695. [Google Scholar] | [Crossref] - Sinwat N, Witoonsatian K, Chumsing S, Suwanwong M, Kankuntod S, Jirawattanapong P. Antimicrobial resistance phenotypes and genotypes of
Salmonella spp. isolated from commercial duck meat production in Thailand and their minimal inhibitory concentration of disinfectants. Microb Drug Resist 2021;27(10):1733-41. [Google Scholar] | [Crossref] - Flores Monter YM, Chaves A, Arellano-Reynoso B, López-Pérez AM, Suzán-Azpiri H, Suzán G. Edaphoclimatic seasonal trends and variations of the
Salmonella spp. infection in northwestern Mexico. Infect Dis Model 2021;6:805-19. [Google Scholar] | [Crossref] - Al-Hadidi SH, Al Mana H, Almoghrabi SZ, El-Obeid T, AlAli WQ, Eltai NO. Retail chicken carcasses as a reservoir of multidrug-resistant
Salmonella . Microb Drug Resist 2022;28(7):824-31. [Google Scholar] | [Crossref] - Mooijman KA. The new ISO 6579-1:a real horizontal standard for detection of
Salmonella , at last!Food Microbiol 2018;71:2-7. [Google Scholar] | [Crossref] - Abd El-Aziz NK, Tartor YH, Gharieb RMA, Erfan AM, Khalifa E, Said MA. Extensive drug-resistant
Salmonella enterica isolated from poultry and humans:prevalence and molecular determinants behind the co-resistance to ciprofloxacin and tigecycline. Front Microbiol 2021;12:738784. [Google Scholar] | [Crossref] - Humphries R, Bobenchik AM, Hindler JA, Schuetz AN. Overview of changes to the Clinical and Laboratory Standards Institute performance standards for antimicrobial susceptibility testing, M100, 31st edition. J Clin Microbiol 2021;59(8):e0021321. [Google Scholar] | [Crossref]
- Bearson SMD.
Salmonella in swine:prevalence, multidrug resistance, and vaccination strategies. Annu Rev Anim Biosci 2022;10:373-93. [Google Scholar] | [Crossref] - Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis:criteria for bacterial strain typing. J Clin Microbiol 1995;33(9):2233-9. [Google Scholar] | [Crossref]
- Chen C, Zhao Y, Han H, Pang B, Zhang J, Yan M. Optimization of pulsed-field gel electrophoresis protocols for
Salmonella Paratyphi A subtyping. Foodborne Pathog Dis 2012;9(4):325-30. [Google Scholar] | [Crossref] - Machado Junior PC, Chung C, Hagerman A. Modeling
Salmonella spread in broiler production:identifying determinants and control strategies. Front Vet Sci 2020;7:564. [Google Scholar] | [Crossref] - Zhao L, Liu G, Tang W, Song X, Zhao X, Wang C. Antimicrobial resistance and genomic characteristics of
Salmonella from broilers in Shandong Province. Front Vet Sci 2023;10:1292401. [Google Scholar] | [Crossref] - Sun T, Liu Y, Qin X, Aspridou Z, Zheng J, Wang X. The prevalence and epidemiology of
Salmonella in retail raw poultry meat in China:a systematic review and meta-analysis. Foods 2021;10(11):2757. [Google Scholar] | [Crossref] - Kim T, Kim Y, Kim H, Moon JS, Chon J, Song K-Y. Prevalence of
Salmonella serotypes isolated from clinical samples in chicken farms and meat in slaughterhouses in South Korea. Poult Sci 2025;104(3):105147. [Google Scholar] | [Crossref] - Roberts T, Rattanavong S, Phommasone K, Chansamouth V, Davong V, Keoluangkhot V. Typhoid in Laos:an 18-year perspective. Am J Trop Med Hyg 2020;102(4):749. [Google Scholar] | [Crossref]
- Hooda Y, Tanmoy AM, Saha SK, Saha S. Genomic surveillance of
Salmonella Paratyphi A:neglected no more?. Open Forum Infect Dis 2023;10((Suppl 1)):S53-7. [Google Scholar] | [Crossref] - Damtew YT, Tong M, Varghese BM, Anikeeva O, Hansen A, Dear K. The impact of temperature on non-typhoidal
Salmonella andCampylobacter infections:an updated systematic review and meta-analysis of epidemiological evidence. EBioMedicine 2024;109:105393. [Google Scholar] | [Crossref] - Buawiratlert T, Tadee P, Chotinun S, Chuammitri P, Kheowsri S, Suddee W. Prevalence and antimicrobial-resistant patterns of non-typhoidal
Salmonella in good agricultural practice certified broiler farms and poultry slaughterhouses in an intensive farming area in upper northern part of Thailand. Vet Integr Sci 2023;21((3)). [Google Scholar] | [Crossref] - Noenchat P, Direksin K, Sornplang P. The phenotypic and genotypic antimicrobial resistance patterns of
Salmonella isolated from chickens and meat at poultry slaughterhouses in Japan and Thailand. Vet World 2023;16(7):1527-33. [Google Scholar] | [Crossref] - Angkititrakul S, Chomvarin C, Chaita T, Kanistanon K, Waethewutajarn S. Epidemiology of antimicrobial resistance in
Salmonella isolated from pork, chicken meat and humans in Thailand. Southeast Asian J Trop Med Public Health 2005;36(6):1510-5. [Google Scholar] | [Crossref] - Sirichote P, Bangtrakulnonth A, Tianmanee K, Unahalekhaka A, Oulai A, Chittaphithakchai P. Serotypes and antimicrobial resistance of
Salmonella enterica ssp in central Thailand, 2001-2006. Southeast Asian J Trop Med Public Health 2010;41(6):1405-15. [Google Scholar] | [Crossref] - Díez-García M, Capita R, Alonso-Calleja C. Influence of serotype on the growth kinetics and the ability to form biofilms of
Salmonella isolates from poultry. Food Microbiol 2012;31(2):173-80. [Google Scholar] | [Crossref] - Salazar JK, Deng K, Tortorello ML, Brandl MT, Wang H, Zhang W. Genes ycfR, sirA and yigG contribute to the surface attachment of
Salmonella enterica Typhimurium and Saintpaul to fresh produce. PLoS One 2013;8(2):e57272. [Google Scholar] | [Crossref] - Phaophu P, Thadtapong N, Wirth SE, Gray AS, Dangsuk S, Ngamwongsatit N. High prevalence of multidrug-resistant
Salmonella enterica in Thailand food markets:insights from complete genome and phenotypic characterization of ESBL-producing strains. Microbiol Spectr 2025;13(2):e0212925. [Google Scholar] | [Crossref] - Lertworapreecha M, Noomee S, Sutthimusik S, Utarapichat B, Tontikapong K. Multidrug-resistant and extended spectrum β-lactamase producing
Salmonella enterica isolated from food animals in Phatthalung, Thailand. Southeast Asian J Trop Med Public Health 2016;47(6):1257-69. [Google Scholar] | [Crossref] - Sripaurya B, Pelyuntha W, Ngasaman R, Ching CL, Guyonnet V, Vongkamjan K. Antibiotic resistance, sequence typing, and virulence gene profiles of
Salmonella enterica isolated from the broiler production chain in southern Thailand. Res Vet Sci 2025;196:105891. [Google Scholar] | [Crossref] - Zhang Z, Suksawat F, Zhang X, Shu X, Angkititrakul S. Clonal and plasmid-mediated dissemination of multidrug-resistant
Salmonella Enteritidis in chicken production, northeastern Thailand. Pathogens 2026;15(1):75. [Google Scholar] | [Crossref] - Tuat CV, Hue PT, Loan NTP, Thuy NT, Hue LT, Giang VN. Antimicrobial resistance pilot surveillance of pigs and chickens in Vietnam, 2017-2019. Front Vet Sci 2021;8:618497. [Google Scholar] | [Crossref]
- Pungpian C, Lee S, Trongjit S, Sinwat N, Angkititrakul S, Prathan R. Colistin resistance and plasmid-mediated mcr genes in
Escherichia coli andSalmonella isolated from pigs, pig carcass and pork in Thailand, Lao PDR and Cambodia border provinces. J Vet Sci 2021;22(5):e68. [Google Scholar] | [Crossref] - Mohammed Jajere S, Hassan L, Zakaria Z, Abu J, Abdul Aziz S. Antibiogram profiles and risk factors for multidrug resistance of
Salmonella enterica recovered from village chickens (Gallus gallus domesticus Linnaeus) and other environmental sources in the central and southern peninsular Malaysia. Antibiotics (Basel) 2020;9(10):701. [Google Scholar] | [Crossref] - Chonsin K, Changkwanyeun R, Siriphap A, Intarapuk A, Prapasawat W, Changkaew K. Prevalence and multidrug resistance of
Salmonella in swine production chain in a central province, Thailand. J Food Prot 2021;84(12):2174-84. [Google Scholar] | [Crossref] - Meunsene D, Eiamsam-Ang T, Patchanee P, Pascoe B, Tadee P. Molecular evidence for cross boundary spread of
Salmonella spp. in meat sold at retail markets in the middle Mekong basin area. PeerJ 2021;9:e11255. [Google Scholar] | [Crossref] - Prasertsee T, Pascoe B, Patchanee P. Colistin resistance and resistance determinants are mobile among
Salmonella enterica isolates from diseased and healthy pigs in Thailand. bioRxiv 2023. [Google Scholar] | [Crossref]