

ABSTRACT
Background and Aim: Native Blackbone chickens (
Materials and Methods: A prospective longitudinal study was conducted between August 2020 and July 2021 on a small-scale Blackbone chicken farm in Phitsanulok province, Thailand. A total of 2,258 samples, comprising 1,755 fecal and 503 meat samples, were collected from chickens aged 1–50 weeks.
Results: The overall prevalence of
Conclusion: Thai native Blackbone chickens harbor non-typhoidal
Keywords: antimicrobial resistance, Blackbone chicken, food safety, genomic characterization, native poultry, non-typhoidal
INTRODUCTION
Non-typhoidal
Human salmonellosis is commonly characterized by mild, self-limiting gastroenteritis with low mortality and often does not require antimicrobial treatment. However, untreated infections may progress to invasive disease, including bacteremia, meningitis, and other extraintestinal manifestations, which can be fatal. Severe salmonellosis has also been associated with an increased risk of colon cancer [5]. Children, elderly individuals, and immunocompromised patients are particularly vulnerable to invasive
Although human salmonellosis is a worldwide disease, severe effects from
The global rise of antimicrobial-resistant
Native chickens are widely raised in underdeveloped and developing countries and play an important role in supporting household livelihoods. In Thailand, diverse native chicken breeds exist, including Blackbone chickens, which are distinguished by their white feathers and black beak, bones, and skin, and their black meat. Consumption of Blackbone chickens has increased due to their perceived health benefits, including antioxidant properties, low cholesterol content, and potential neuroprotective effects demonstrated in experimental models [14–16].
Several pathogenic bacteria, including
Despite extensive research on
Despite the well-documented role of commercial poultry systems in the epidemiology of non-typhoidal
Moreover, information on the AMR and virulence potential of
The present study was designed to address these knowledge gaps by comprehensively characterizing non-typhoidal
By integrating longitudinal sampling with phenotypic and genomic analyses, this study aimed to generate baseline molecular epidemiological data on
MATERIALS AND METHODS
Ethical approval
This study was conducted in full compliance with institutional, national, and international guidelines governing the ethical use of animals in research. The study protocol was reviewed and approved by the Animal Care and Use Committee of Chulalongkorn University under animal use protocol number 2031019 (approval date: July 22, 2020). All experimental and sampling procedures adhered strictly to the principles of animal welfare, including the minimization of stress, discomfort, and disturbance to animals during routine farm activities and sample collection.
Fecal sampling was non-invasive and involved the collection of freshly voided feces from the ground, thereby avoiding direct handling or restraint of birds. Meat samples were obtained from birds slaughtered as part of standard farm management and commercial processing practices, and no animals were euthanized specifically for research purposes. Prior to commencement of the study, verbal informed consent was obtained from the farm owner for on-farm observations, interviews, and sample collection.
All laboratory procedures involving
The study did not involve human participants, human biological samples, or the collection of personal or identifiable data. Therefore, approval from a human research ethics committee was not required. Overall, the study was designed and implemented to ensure ethical integrity, biosafety, and responsible conduct of research, consistent with the principles of the One Health framework.
Study period and location
The study was carried out over a 12-month period from August 2020 to July 2021 at a Blackbone chicken farm located in Phitsanulok province (16°49′N, 100°15′E), approximately 377 km north of Bangkok in the lower northern region of Thailand. The area has a tropical climate characterized by a hot, dry season from November to April and a hot, wet season from May to October. Mean temperatures range from 27.1°C during the dry season to 28.6°C during the wet season, with an average annual humidity of 72% and an annual rainfall of approximately 1662 mm, predominantly occurring during the wet season.
Study design and farm management
A prospective longitudinal surveillance study was conducted to evaluate the prevalence and antimicrobial resistance of
Sample size calculation and sampling scheme
Sample size determination was performed using the Epitools online software (http://epitools.ausvet. com.au). As the true prevalence of
Fecal and meat sample collection
Fresh fecal samples were collected weekly from the ground from chickens aged 1–5 weeks using sterile spoons and placed into disposable containers. Subsequent sampling was conducted at 7, 10, 14, 16, 22, 28, 40, and 50 weeks of age. Care was taken to avoid feces that had come into contact with the ground surface.
Meat samples were collected from all birds slaughtered at 7, 10, and 16 weeks of age. A comparable number of meat samples was randomly collected when the remaining birds were slaughtered at the end of the production cycle (50 weeks of age). Chicken carcasses were individually placed in sterile plastic bags, transported on ice, and processed within 24 h of collection.
Isolation and culture of Salmonella spp.
For fecal samples, 1 g of each sample was enriched in 9 mL buffered peptone water (BPW) (Difco™, MD, USA) and incubated at 37°C for 24 h. For meat samples, 25 g of tissue was placed in a sterile bag containing 225 mL BPW, homogenized for 2 min using a stomacher, and incubated at 37°C for 24 h. Subsequently, 0.1 mL of enriched BPW was transferred into 9.9 mL Rappaport–Vassilliadis soya peptone (RVS) broth and incubated at 42°C ± 1°C for 24 h, while 1 mL of BPW was transferred into 9 mL tetrathionate (TT) broth (Oxoid, Hampshire, UK) and incubated at 37°C ± 1°C for 24 h. A loopful from each incubated broth was streaked onto Xylose Lysine Deoxycholate (XLD) agar (Oxoid) and incubated overnight at 37°C.
Biochemical and molecular confirmation of Salmonella spp.
Presumptive
All confirmed
Serovar identification
Initial serogrouping of isolates was performed by slide agglutination using
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed using the disk diffusion method in accordance with Clinical and Laboratory Standards Institute (CLSI) guidelines [27]. Isolates were grown overnight on Trypticase soy agar at 37°C. Three to five colonies were adjusted to a 0.5 McFarland standard (1.5 × 108 CFU/mL) in normal saline and evenly spread onto Mueller–Hinton agar (MHA) plates (4 mm thickness, pH 7.2–7.4; Oxoid). Antibiotic disks were applied, and plates were incubated at 35 ± 2°C for 16–18 h. Inhibition zone diameters were measured and interpreted according to CLSI criteria (CLSI M100, 30th ed., 2020) [27].
The antimicrobial agents tested included β-lactams (ampicillin 10 µg, cefazolin 30 µg, cefuroxime 30 µg, cefotaxime 30 µg, and ceftazidime 30 µg), aminoglycosides (streptomycin 10 µg and gentamicin 10 µg), tetracycline (doxycycline 30 µg), quinolone (ciprofloxacin 5 µg), phenicol (chloramphenicol 30 µg), folate pathway antagonist (trimethoprim/sulfamethoxazole 25 µg), fosfomycin 200 µg, and nitrofurantoin 300 µg (Oxoid). All disks were stored at 2°C–4°C in sealed containers protected from light according to the manufacturer’s instructions.
Colistin minimum inhibitory concentration (MIC) determination
Colistin susceptibility was determined by broth microdilution to establish the MIC following CLSI guidelines [27]. Twofold serial dilutions of colistin (Sigma-Aldrich, MO, USA) were prepared in 200 µL cation-adjusted Mueller–Hinton broth (Oxoid) in microtiter plates. Bacterial suspensions were added to achieve a final inoculum of 5 × 105 CFU/mL per well, and plates were incubated at 35 ± 2°C for 18–20 h. The MIC was defined as the lowest concentration inhibiting visible growth. Isolates with MIC ≥ 4 µg/mL were classified as colistin resistant (Table S2) (CLSI M100, 30th ed., 2020) [27].
Genomic DNA extraction and WGS
Six
Genome assembly, annotation, and in silico analyses
All bioinformatic analyses were conducted using default parameters unless otherwise specified. Raw sequencing reads were assembled using Unicycler v0.4.8 [29] via the Bacterial and Viral Bioinformatics Resource Center (https://www.bv-brc.org). Genome annotation was performed using Prokka v1.14.6 [30]. Species identification was confirmed by average nucleotide identity analysis based on ANIb using the JSpeciesWS online server [31]. Multilocus sequence typing was conducted using MLST v2.0.9 [32], and antimicrobial resistance genes were identified using ResFinder v4.6.0 (identity ≥90%, coverage ≥80%) [33] through the Center for Genomic Epidemiology (https://www.genomicepidemiology.org). MobileElementFinder v1.0.3 was used to identify plasmid replicon types and the presence of the colicin Ib gene (cib) [34]. Virulence-associated genes were identified using the virulence factor analyzer based on the Virulence Factor Database 2022 (VFDB) with default settings [35].
Phylogenetic analysis based on core genome SNPs
Phylogenetic relationships among
Statistical analysis
Descriptive statistics were used to summarize prevalence and AMR data. Differences in detection rates between fecal and meat samples and among sampling periods were analyzed using chi-square or Fisher’s exact tests. Statistical analyses were performed using SPSS version 17.0, with p < 0.05 considered statistically significant.
RESULTS
Prevalence of Salmonella spp. in Blackbone chickens
The overall prevalence of
Table 1.
| Age (week) | Feces | Meat | ||||
|---|---|---|---|---|---|---|
|
|
| |||||
| No. | Serovars | No. | Serovars | |||
| 1 | 155 | 0 | – | – | – | – |
| 2 | 155 | 0 | – | – | – | – |
| 3 | 155 | 8 (5.2) (2.3–9.9) | Bovismorbificans (n = 7) Weltevreden (n = 1) | – | – | – |
| 4 | 125 | 4 (3.2) (0.9–8.0) | Bovismorbificans (n = 4) | – | – | – |
| 5 | 125 | 9 (7.2) (3.3–13.2) | Bovismorbificans (n = 6) Weltevreden (n = 2) Corvallis (n = 1) | – | – | – |
| 7 | 130 | 26 (20.0) (13.5–27.9) | Bovismorbificans (n = 15) Corvallis (n = 6) Weltevreden (n = 2) Stanley (n = 2) Typhimurium (n = 1) | 125 | 27 (21.6) (14.7–29.8) | Bovismorbificans (n = 20) Weltevreden (n = 5) Stanley (n = 2) |
| 10 | 130 | 14 (10.8) (6.0–17.4) | Corvallis rats (n = 13) Weltevreden (n = 1) | 126 | 11 (8.7) (4.4–15.1) | Bovismorbificans (n = 9) Weltevreden (n = 1) Stanley (n = 1) |
| 14 | 130 | 8 (6.2) (2.7–11.8) | Bovismorbificans (n = 8) | – | – | – |
| 16 | 130 | 3 (2.3) (0.5–6.6) | Bovismorbificans (n = 3) | 126 | 20 (15.9) (10.0–23.4) | Bovismorbificans (n = 17) Kentucky (n = 2) Corvallis (n = 1) |
| 22 | 130 | 1 (0.8) (0.1–4.2) | Corvallis (n = 1) | – | – | – |
| 28 | 130 | 3 (2.3) (0.5–6.6) | Corvallis (n = 3) | – | – | – |
| 40 | 130 | 2 (1.5) (0.2–5.4) | Corvallis (n = 1) Serogroup C (n = 1) | – | – | – |
| 50 | 130 | 4 (3.1) (0.8–7.7) | Corvallis (n = 3) Weltevreden (n = 1) | 126 | 2 (1.6) (0.2–5.6) | Bovismorbificans (n = 2) |
| Total | 1,755 | 82 (4.7) (3.1–5.8) | Bovismorbificans (n = 43) Corvallis (n = 28) Weltevreden (n = 7) Stanley (n = 2) Typhimurium (n = 1) Serogroup C (n = 1) | 503 | 60 (11.9) (9.2–15.1) | Bovismorbificans (n = 48) Weltevreden (n = 6) Stanley test (n = 3) Kentucky (n = 2) Corvallis (n = 1) |
a Serovar identification was untypeable; however, typing performed by agglutination with
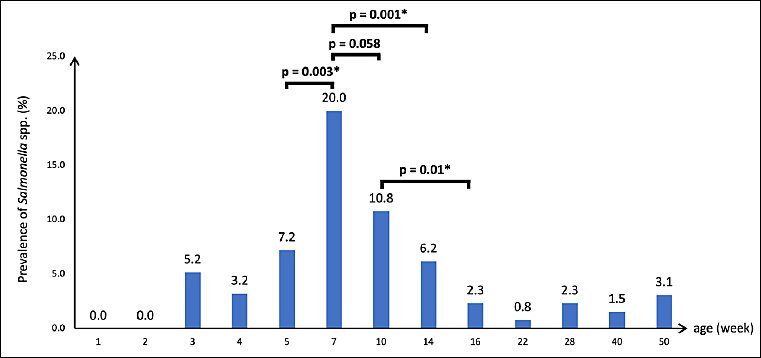
Figure 1.
In meat samples,
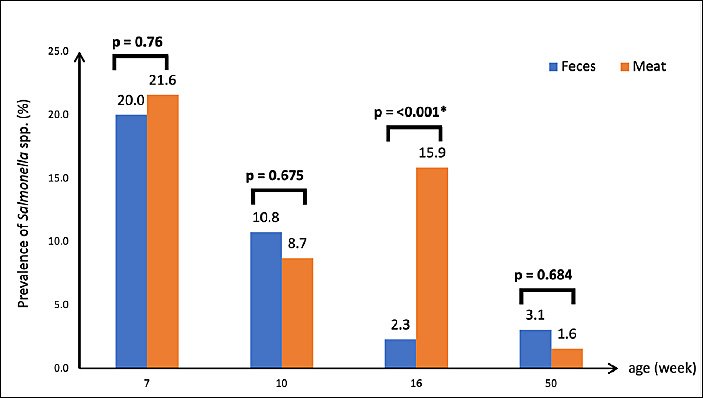
Figure 2.
Serovar distribution of Salmonella spp.
Serovar analysis identified six distinct
Table 2. Antibiotic resistance profiles of
| Antibiotic resistance profile | Number of antibiotic classes | Number of isolates, n (%) (95% CI) | Serovar (n) |
|---|---|---|---|
| 1. KZ-S-DO-F | 4 | 1 (0.7) (0.02–3.9) | Bovismorbificans (n = 1) |
| 2. AMP-KZ-S-CN | 3 | 1 (0.7) (0.02–3.9) | Bovismorbificans (n = 1) |
| 3. AMP-KZ-S | 2 | 3 (2.1) (0.4–6.0) | Corvallis (n = 3) |
| 4. AMP-KZ-DO | 2 | 4 (2.8) (0.8–7.1) | Bovismorbificans (n = 3), Weltevreden (n = 1) |
| 5. AMP-KZ | 1 | 3 (2.1) (0.4–6.0) | Corvallis (n = 2), Weltevreden (n = 1) |
| 6. AMP-S | 2 | 1 (0.7) (0.02–3.9) | Corvallis (n = 1) |
| 7. KZ | 1 | 1 (0.7) (0.02–3.9) | Bovismorbificans (n = 1) |
| 8. S | 1 | 60 (42.3) (34.0–50.8) | Bovismorbificans (n = 39), Corvallis (n = 18), Stanley (n = 1), Typhimurium (n = 1), Serogroup C (n = 1) |
| 9. TMP/SMX | 1 | 1 (0.7) (0.02–3.9) | Stanley (n = 1) |
| Subtotal (resistant isolates) | 75 (52.8) (44.3–61.2) | ||
| Susceptible to 13 antibiotics tested | 67 (47.2) (38.8–55.7) | ||
| Total | 142 | ||
AMP = Ampicillin, CN = Gentamicin, DO = Doxycycline, F = Nitrofurantoin, KZ = Cefazolin, S = Streptomycin, TMP/SMX = Trimethoprim/ sulfamethoxazole.
a Isolates were defined as multidrug-resistant.
Antimicrobial susceptibility patterns
All

Figure 3.
Nine distinct resistance profiles were identified, with most resistant isolates (42.3%, 60/142) displaying resistance to streptomycin alone (Table 2). MDR was rare, occurring in only 1.4% (n = 2) of
WGS and genomic features
Six
Average nucleotide identity analysis confirmed all isolates as
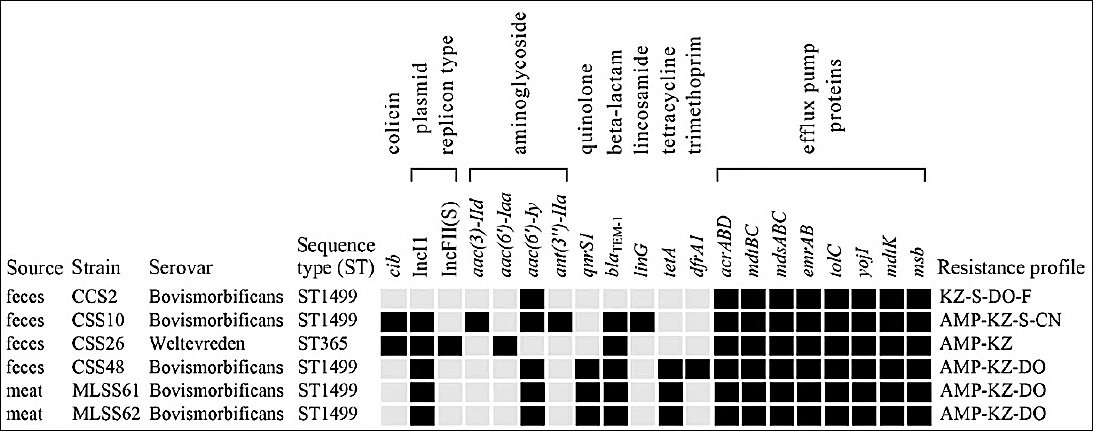
Figure 4. Sequence types, antibiotic-resistant genes, plasmid replicon types, and colicin Ib (
ARGs and plasmid profiles
Genomic analysis revealed ARGs associated with six antibiotic classes (Figure 4). All isolates harbored aminoglycoside resistance genes, including
Plasmid analysis showed that five isolates carried an IncI1 plasmid. Two of these isolates (one
Virulence gene profiles
Virulence factor analysis identified between 158 and 162 virulence-associated genes per isolate (Figure 5), with 79.6% of genes conserved across all strains.
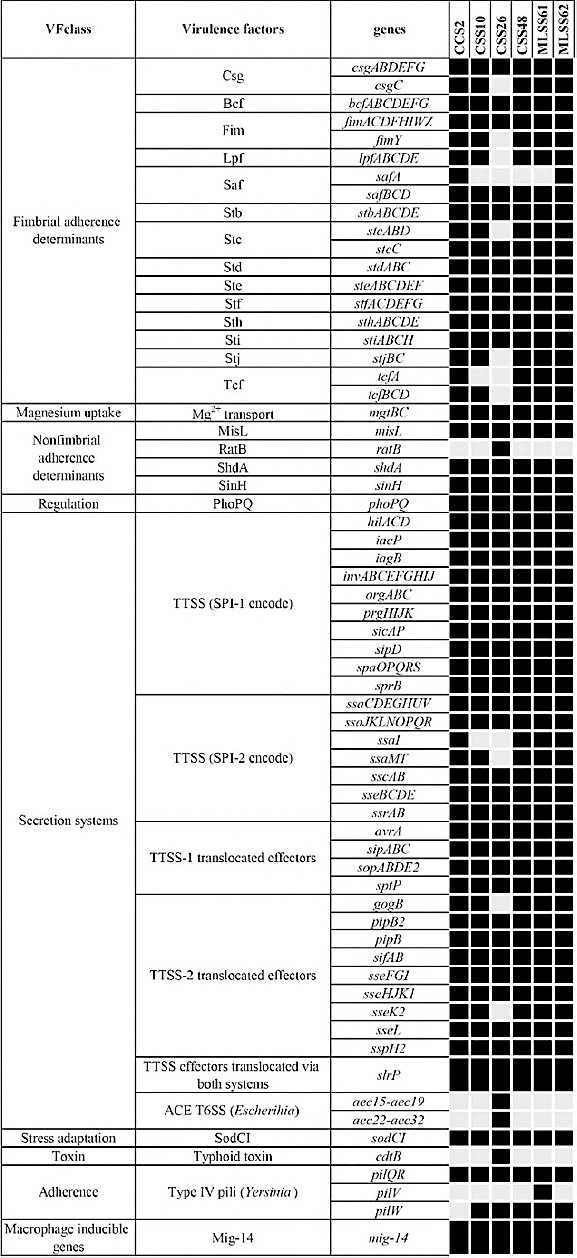
Figure 5. Virulence factor-associated genes in Blackbone chicken isolates of selected
Notably,
Phylogenetic relationships
Core genome single-nucleotide polymorphism(cgSNP)-based phylogenetic analysis revealed that
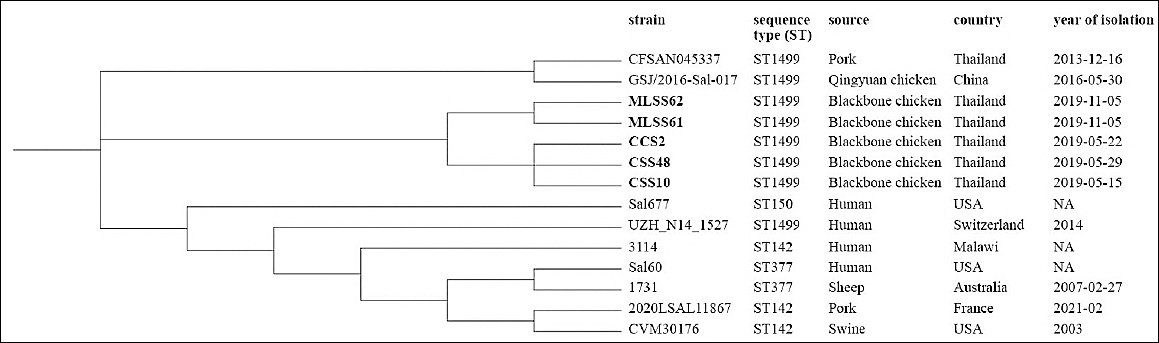
Figure 6. Core genome single-nucleotide polymorphism-based phylogenetic tree of
Phylogenetic analysis of

Figure 7. Core genome single-nucleotide polymorphism-based phylogenetic tree of
DISCUSSION
Prevalence of Salmonella spp. in Blackbone chickens
Regarding the prevalence of
Age-related fecal shedding and seasonal influence
At the farm level,
Then,
Distribution of serotypes and epidemiological significance
The distribution of serovars of
As opposed to previous studies, our study demonstrated that
Although several
In Thailand,
Trends in antibiotic resistance and public health implications
The incidence of antibiotic-resistant
Compared with the
Several
Genomic findings and virulence-associated characteristics
Previous studies have shown that the common
All isolates harbored a limited number of ARGs, consistent with the finding that most remained susceptible to antibiotics (Table 2). The presence of
Plasmids are important vehicles for the conjugative transfer of genes. In this study, five isolates carried an IncI1 plasmid, a broad-host-range plasmid that typically harbors multiple ARGs. This is not surprising, as the IncI1 plasmid is frequently found in Enterobacterales of food-animal origin, including
Chromosome-encoded SPI-1 and SPI-2 are necessary for
Interestingly, we identified a single
The CSS26 strain lacked some fimbrial adhesin genes as well as the
The chromosome-encoded ACE T6SS gene cluster, which has been reported to be associated with gut bacteria killing and outcompeting [71], is present in CSS26. Consequently, the presence of
Note that all 6
Phylogenetic relationships
The cgSNP analysis revealed that five
In contrast, cgSNP analysis revealed that
CONCLUSION
This longitudinal investigation provides comprehensive evidence on the occurrence, AMR, and genomic characteristics of non-typhoidal
The detection of potentially invasive and zoonotic
A major strength of this study lies in its prospective longitudinal design, which enabled age-specific assessment of
Several limitations should be acknowledged. The study was conducted on a single farm, which may limit generalizability to other native poultry systems. Environmental samples, including water, feed, litter, and slaughterhouse surfaces, were not collected, restricting the ability to identify transmission pathways. In addition, WGS was performed on a limited number of isolates, which may underestimate the genetic diversity of circulating
Future studies should incorporate multi-farm sampling across different geographic regions and include comprehensive environmental and slaughterhouse surveillance to better elucidate transmission dynamics. Expanded WGS of larger numbers of isolates is warranted to monitor the emergence of resistance, virulence evolution, and interspecies transmission. Longitudinal One Health studies linking poultry, human, and environmental isolates would further strengthen risk assessment and inform evidence-based control strategies.
In conclusion, Thai native Blackbone chickens harbor non-typhoidal
DATA AVAILABILITY
The associated metadata (e.g., serovar, resistance phenotype, and sample source) are included in this study. The raw sequences of
AUTHORS’ CONTRIBUTIONS
KA: Methodology, fieldwork, performing experiments, data collection, curation, and analysis. AK and UT: Methodology and performing experiments. PRN: Fieldwork, data analysis and interpretation, and drafting the manuscript. PRN and TL: Conceptualization, supervision, methodology, data analysis and interpretation, and reviewing and editing the manuscript. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
This work was supported by Naresuan University (NU) and the National Science Research and Innovation Fund (NSRF), Grant No. R2566B042, and partially funded by the National Science and Technology Development Agency (NSTDA), Thailand (FDA-CO-2561-6029-TH). K. Assawatheptawee was supported by the Royal Golden Jubilee-PhD program from Thailand Science Research and Innovation (PHD/0123/2560).
REFERENCES
- Galán-Relaño Á, Valero Díaz A, Huerta Lorenzo B, Gómez-Gascón L, Mena Rodríguez MÁ, Carrasco Jiménez E.
Salmonella and salmonellosis:an update on public health implications and control strategies. Animals 2023;13(23):3666. [Google Scholar] | [Crossref] - Knoblauch AM, Bratschi MW, Zuske MK, Althaus D, Stephan R, Hächler H. Cross-border outbreak of
Salmonella enterica ssp enterica serovar Bovismorbificans:multiple approaches for an outbreak investigation in Germany and Switzerland. Swiss Med Wkly 2015;145:w14182. [Google Scholar] | [Crossref] - Kongsanan P, Angkititrakul S, Kiddee A, Tribuddharat C. Spread of antimicrobial-resistant
Salmonella from poultry to humans in Thailand. Jpn J Infect Dis 2021;74(3):220-227. [Google Scholar] | [Crossref] - Politi L, Mellou K, Chrysostomou A, Mandilara G, Spiliopoulou I, Theofilou A. A community waterborne
Salmonella Bovismorbificans outbreak in Greece. Int J Environ Res Public Health 2024;21(2):167. [Google Scholar] | [Crossref] - Mughini-Gras L, Schaapveld M, Kramers J, Mooij S, Neefjes-Borst EA, van Pelt W. Increased colon cancer risk after severe
Salmonella infection. PLoS One 2018;13(1):e0189721. [Google Scholar] | [Crossref] - Burden of foodborne diseases in the South-East Asia Region. 2016. Accessed 7 Jul 2025. [Available from] | [Google Scholar]
- WHO bacterial priority pathogens list 2024:bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance. 2024. Accessed 7 Jul 2025. [Available from] | [Google Scholar]
- Tribuddharat C, Pongpech P, Srifuengfung S, Meethai C. Prevalence and drug susceptibility of
Salmonella isolated from patients in Bangkok, Thailand. J Glob Antimicrob Resist 2016;6:162-164. [Google Scholar] | [Crossref] - Hengkrawit K, Tangjade C. Prevalence and trends in antimicrobial susceptibility patterns of multidrug-resistant non-typhoidal
Salmonella in central Thailand, 2012–2019. Infect Drug Resist 2022;15:1305-1315. [Google Scholar] | [Crossref] - Buddhasiri S, Sukjoi C, Tantibhadrasapa A, Mongkolkarvin P, Boonpan P, Pattanadecha T. Clinical characteristics, antimicrobial resistance, virulence genes and multilocus sequence typing of non-typhoidal
Salmonella serovar Typhimurium and Enteritidis strains isolated from patients in Chiang Mai, Thailand. Microorganisms 2023;11(10):2425. [Google Scholar] | [Crossref] - Phongaran D, Khang-Air S, Angkititrakul S. Molecular epidemiology and antimicrobial resistance of
Salmonella isolates from broilers and pigs in Thailand. Vet World 2019;12(8):1311-1318. [Google Scholar] | [Crossref] - Noenchat P, Direksin K, Sornplang P. Phenotypic and genotypic antimicrobial resistance patterns of
Salmonella isolated from chickens and meat at poultry slaughterhouses in Japan and Thailand. Vet World 2023;16(7):1527-1533. [Google Scholar] | [Crossref] - Sinwat N, Witoonsatian K, Chumsing S, Suwanwong M, Kankuntod S, Jirawattanapong P. Antimicrobial resistance phenotypes and genotypes of
Salmonella spp isolated from commercial duck meat production in Thailand and their minimal inhibitory concentration of disinfectants. Microb Drug Resist 2021;27(12):1733-1741. [Google Scholar] | [Crossref] - Zhang Z, Li G, Wei Y, Feng Z, Fang L, Li M. In vitro immunomodulatory and antioxidant effects of oligopeptides and four characteristic peptides in blackbone silky fowl (Gallus gallus domesticus). J Food Biochem 2022;46(12):e14469. [Google Scholar] | [Crossref]
- Jaturasitha S, Srikanchai T, Kreuzer M, Wicke M. Differences in carcass and meat characteristics between chicken indigenous to northern Thailand (black-boned and Thai native) and imported extensive breeds (Bresse and Rhode Island Red). Poult Sci 2008;87(1):160-169. [Google Scholar] | [Crossref]
- Klinprathap K, Kasekarn W, Sattayakawee S, Phothi T, Khongsombat O. Effects of blackbone chicken on learning and memory in oxonic-induced hyperuricemia male rats. Funct Food Sci 2025;5(3):97-112. [Google Scholar] | [Crossref]
- Balala LM, Mendoza BC, Baldrias LR, Masangkay JS. Detection of multidrug-resistant
Salmonella in native chickens by culture, polymerase chain reaction, and enzyme-linked immunosorbent assay. Asia Pac J Sci Technol 2024;30(1):APST-30. [Google Scholar] | [Crossref] - Jajere SM, Hassan L, Abdul-Aziz S, Zakaria Z, Abu J, Nordin F.
Salmonella in native village chickens (Gallus domesticus):prevalence and risk factors from farms in south-central Peninsular Malaysia. Poult Sci 2019;98(11):5961-5970. [Google Scholar] | [Crossref] - Boraei-Nezhad G, Saadati D, Jahantigh M, Saadat-Jou S. Prevalence of
Salmonella infection in village chickens and tetracycline resistance genes in isolates from the Sistan region, Iran. Braz J Microbiol 2023;54(3):2375-2382. [Google Scholar] | [Crossref] - Cui L, Liu Q, Jiang Z, Song Y, Yi S, Qiu J. Characteristics of
Salmonella from Chinese native chicken breeds fed conventional or antibiotic-free diets. Front Vet Sci 2021;8:607491. [Google Scholar] | [Crossref] - Laidlow TA, Stafford R, Jennison AV, Bell R, Graham R, Graham T. A multi-jurisdictional outbreak of
Salmonella Typhimurium infections linked to backyard poultry, Australia, 2020. Zoonoses Public Health 2022;69(7):835-842. [Google Scholar] | [Crossref] - Stapleton GS, Habrun C, Nemechek K, Gollarza L, Ellison Z, Tolar B. Multistate outbreaks of salmonellosis linked to contact with backyard poultry, United States, 2015–2022. Zoonoses Public Health 2024;71(6):708-722. [Google Scholar] | [Crossref]
- Nichols M, Gollarza L, Palacios A, Stapleton GS, Basler C, Hoff C.
Salmonella illness outbreaks linked to backyard poultry purchasing during the COVID-19 pandemic, United States, 2020. Epidemiol Infect 2021;149:e234. [Google Scholar] | [Crossref] - Kunjara C, Kupopan S, Srisuparbh K, Santivatr D. Study on
Salmonella carrier rate in native chickens. Agric Nat Resour 1987;21(1):46-53. [Google Scholar] | [Crossref] - Assawatheptawee K, Punyadi P, Luangtongkum T, Niumsup PR. Longitudinal fecal shedding patterns and characterization of
Salmonella enterica andmcr -positiveEscherichia coli in meat-type ducks raised in an open-house system. Poult Sci 2022;101(10):102090. [Google Scholar] | [Crossref] - Rahn K, De Grandis SA, Clarke RC, McEwen SA, Galán JE, Ginocchio C. Amplification of an invA gene sequence of
Salmonella Typhimurium by polymerase chain reaction as a specific method of detection ofSalmonella . Mol Cell Probes 1992;6((4)):271-279. [Google Scholar] | [Crossref] - Performance standards for antimicrobial susceptibility testing. Wayne (PA): CLSI; 2020. [Google Scholar]
- Falagas ME, Karageorgopoulos DE. Pandrug resistance, extensive drug resistance and multidrug resistance among Gram-negative bacilli:need for international harmonization in terminology. Clin Infect Dis 2008;46(7):1121-1122. [Google Scholar] | [Crossref]
- Wick RR, Judd LM, Gorrie CL, Holt KE. Unicycler:resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol 2017;13(6):e1005595. [Google Scholar] | [Crossref]
- Seemann T. Prokka:rapid prokaryotic genome annotation. Bioinformatics 2014;30(14):2068-2069. [Google Scholar] | [Crossref]
- Richter M, Rosselló-Móra R, Glöckner FO, Peplies J. JSpeciesWS:a web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016;32:929-931. [Google Scholar] | [Crossref]
- Larsen MV, Cosentino S, Rasmussen S, Friis C, Hasman H, Marvig RL. Multilocus sequence typing of total-genome-sequenced bacteria. J Clin Microbiol 2012;50(4):1355-1361. [Google Scholar] | [Crossref]
- Bortolaia V, Kaas RS, Ruppe E, Roberts MC, Schwarz S, Cattoir V. ResFinder 4.0 for predictions of phenotypes from genotypes. J Antimicrob Chemother 2020;75(12):3491-3500. [Google Scholar] | [Crossref]
- Johansson MHK, Bortolaia V, Tansirichaiya S, Aarestrup FM, Roberts AP, Petersen TN. Detection of mobile genetic elements associated with antibiotic resistance in
Salmonella enterica using MobileElementFinder. J Antimicrob Chemother 2021;76(1):101-109. [Google Scholar] | [Crossref] - Liu B, Zheng D, Zhou S, Chen L, Yang J. VFDB 2022:a general classification scheme for bacterial virulence factors. Nucleic Acids Res 2022;50((D1)):D912-D917. [Google Scholar] | [Crossref]
- Page AJ, Cummins CA, Hunt M, Wong VK, Reuter S, Holden MTG. Roary:rapid large-scale prokaryote pan-genome analysis. Bioinformatics 2015;31(22):3691-3693. [Google Scholar] | [Crossref]
- Kaas RS, Leekitcharoenphon P, Aarestrup FM, Lund O. Solving the problem of comparing whole bacterial genomes across different sequencing platforms. PLoS One 2014;9(8):e104984. [Google Scholar] | [Crossref]
- Letunic I, Bork P. Interactive Tree of Life (iTOL) v6:recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res 2024;52((W1)):W78-W82. [Google Scholar] | [Crossref]
- Wang J, Wu H, Song M, Li F, Zhu J, Xi M. Prevalence and quantitative detection of
Salmonella in retail raw chicken in Shaanxi, China. J Food Prot 2013;76(11):1958-1962. [Google Scholar] | [Crossref] - Boonprasert N, Nuanualsuwan S, Pulsrikarn C, Pornaem S, Chokesajjawatee N. Sources and dissemination of
Salmonella spp. in an integrated broiler meat production system. Thai J Vet Med 2014;44((1)):117-124. [Google Scholar] | [Crossref] - Buawiratlert T, Tadee P, Chotinun S, Chuammitri P, Kheowsri S, Suddee W. Prevalence and antimicrobial-resistant patterns of non-typhoidal
Salmonella in GAP-certified broiler farms and poultry slaughterhouses in northern Thailand. Vet Integr Sci 2023;21((3)). [Google Scholar] | [Crossref] - Khazali D, Ehsani A, Masoudi AA, Torshizi MAK, Torshizi RV, Davoodi P. Comparison of immune-related responses of several slow-growing indigenous chickens with a fast-growing broiler in Iran. Vet Med Sci 2025;11(2):e70288. [Google Scholar] | [Crossref]
- Drauch V, Mitra T, Liebhart D, Hess M, Hess C. Infection dynamics of
Salmonella Infantis vary considerably between chicken lines. Avian Pathol 2022;51(6):561-573. [Google Scholar] | [Crossref] - da Silva Teixeira M, Lages DH, Alves VV, da Silva Martins NR, de Freitas Neto OC. Assessment of maternal immunity against
Salmonella enterica serovar Heidelberg in progeny of broiler breeders vaccinated with bacterins. Avian Pathol 2022;51(2):197-205. [Google Scholar] | [Crossref] - Xaymountry S, Chukanhom K, Klangair S, Jiwakanon J, Angkititrakul S. Seasonal distribution and antimicrobial resistance of
Salmonella isolated from pork, beef and chicken meat in Vientiane Capital, Lao PDR. Thai J Vet Med 2022;52((1)):57-62. [Google Scholar] | [Crossref] - Nair S, Farzan A, O'Sullivan TL, Friendship RM. Time course of
Salmonella shedding and antibody response in naturally infected pigs during the grower–finisher stage. Can J Vet Res 2018;82((2)):139-145. [Google Scholar] | [Crossref] - Gast RK, Guraya R, Jones DR, Anderson KE. Persistence of fecal shedding of
Salmonella Enteritidis by experimentally infected laying hens. Poult Sci 2015;94(7):1650-1656. [Google Scholar] | [Crossref] - Morris VK, Carrique-Mas JJ, Mueller-Doblies D, Davies RH, Wales AD, Allen VM. Longitudinal study of
Salmonella shedding by commercial turkeys during rearing and fattening. Br Poult Sci 2015;56(1):48-57. [Google Scholar] | [Crossref] - Saengthongpinit C, Nane-Siri D, Aparachita P, Apiwannarat P, Buakhao P, Bowornnantiwath W. Longitudinal study of
Salmonella andCampylobacter species in laying duckling flocks in Thailand. Thai J Vet Med 2014;44((3)):355-361. [Google Scholar] | [Crossref] - Toyting J, Nuanmuang N, Utrarachkij F, Supha N, Thongpanich Y, Leekitcharoenphon P. Genomic analysis of
Salmonella isolated from canal water in Bangkok, Thailand. Microbiol Spectr 2024;12(5):e0421623. [Google Scholar] | [Crossref] - Foley SL, Nayak R, Hanning IB, Johnson TJ, Han J, Ricke SC. Population dynamics of
Salmonella enterica serotypes in commercial egg and poultry production. Appl Environ Microbiol 2011;77(13):4273-4279. [Google Scholar] | [Crossref] - Multistate outbreak of
Salmonella serotype Bovismorbificans infections associated with hummus and tahini—United States, 2011. MMWR Morb Mortal Wkly Rep 2012;61(46):944-947. [Google Scholar] | [Crossref] - Brandwagt D, van den Wijngaard C, Tulen AD, Mulder AC, Hofhuis A, Jacobs R. Outbreak of
Salmonella Bovismorbificans associated with uncooked ham products, the Netherlands, 2016–2017. Euro Surveill 2018;23(1):17-00335. [Google Scholar] | [Crossref] - Pardos de la Gandara M, Fournet N, Bonifait L, Lefèvre S, Chemaly M, Grastilleur C. Countrywide multi-serotype outbreak of
Salmonella Bovismorbificans ST142 and monophasicSalmonella Typhimurium ST34 in France, 2020–2021. Euro Surveill 2023;28(2):2200123. [Google Scholar] | [Crossref] - Bronowski C, Fookes MC, Gilderthorp R, Ashelford KE, Harris SR, Phiri A. Genomic characterisation of invasive non-typhoidal
Salmonella enterica serovar Bovismorbificans from Malawi. PLoS Negl Trop Dis 2013;7(11):e2557. [Google Scholar] | [Crossref] - Anuchatkitcharoen C, Numee S, Bender J, Awaiwanont N, Intanon M. Prevalence and antimicrobial resistance of
Salmonella isolated from backyard pigs in Chiang Mai, Thailand. Vet Integr Sci 2020;18(3):1-12. [Google Scholar] | [Crossref] - Percentage of susceptible organisms isolated from all specimens, 68 hospitals, January–December 2022. Accessed 7 Jul 2025. [Available from] | [Google Scholar]
- Van Meervenne E, Van Coillie E, Kerckhof FM, Devlieghere F, Herman L, De Gelder LS. Strain-specific transfer of antibiotic resistance from an environmental plasmid to foodborne pathogens. J Biomed Biotechnol 2012;2012:834598. [Google Scholar] | [Crossref]
- Gopinath GR, Jang H, Beaubrun JJ, Gangiredla J, Mammel MK, Müller A. Phylogenomic analysis of
Salmonella enterica serovar Bovismorbificans from clinical and food samples. Microorganisms 2022;10(6):1199. [Google Scholar] | [Crossref] - Zuo H, Yang Y, Su M, Huang W, Wang J, Lei G. Comparative genomic and antimicrobial resistance profiles of
Salmonella strains from pork and human sources in Sichuan, China. Front Microbiol 2025;16:1515576. [Google Scholar] | [Crossref] - Grossman TH. Tetracycline antibiotics and resistance. Cold Spring Harb Perspect Med 2016;6((4)):a025387. [Google Scholar] | [Crossref]
- Deekshit VK, Srikumar S. “To be, or not to be”:the dilemma of silent antimicrobial resistance genes in bacteria. J Appl Microbiol 2022;133(5):2902-2914. [Google Scholar] | [Crossref]
- Foley SL, Kaldhone PR, Ricke SC, Han J. IncI1 plasmids:genetics, biology and public health relevance. Microbiol Mol Biol Rev 2021;85(2):e00031-20. [Google Scholar] | [Crossref]
- Dos Santos AMP, Ferrari RG, Conte-Junior CA. Virulence factors in
Salmonella Typhimurium:the sagacity of a bacterium. Curr Microbiol 2019;76(6):762-773. [Google Scholar] | [Crossref] - Zhang J, Peng Z, Chen K, Zhan Z, Shen H, Feng S. Genomic characterization of
Salmonella enterica serovar Weltevreden associated with human diarrhea. Microbiol Spectr 2023;11(1):e0354222. [Google Scholar] | [Crossref] - Duong VT, The HC, Nhu TDH, Tuyen HT, Campbell JI, Minh PV. Genomic serotyping, clinical manifestations and antimicrobial resistance of nontyphoidal
Salmonella in children in Vietnam. J Clin Microbiol 2020;58(12):e01465-20. [Google Scholar] | [Crossref] - Meunsene D, Eiamsam-Ang T, Patchanee P, Pascoe B, Tadee P. Molecular evidence for cross-boundary spread of
Salmonella spp. in retail meat in the Mekong basin. PeerJ 2021;9:e11255. [Google Scholar] | [Crossref] - Tadee P, Patchanee P, Pascoe B, Sheppard SK, Meunsene D, Buawiratlert T. Occurrence and sequence types of antimicrobial-resistant
Salmonella spp. in antibiotic-free pig farms in northern Thailand. Thai J Vet Med 2021;51((2)):311-319. [Google Scholar] | [Crossref] - van der Velden AW, Bäumler AJ, Tsolis RM, Heffron F. Multiple fimbrial adhesins are required for full virulence of
Salmonella Typhimurium in mice. Infect Immun 1998;66(6):2803-2808. [Google Scholar] | [Crossref] - Rakov AV, Mastriani E, Liu SL, Schifferli DM. Association of
Salmonella virulence factor alleles with intestinal and invasive serovars. BMC Genomics 2019;20(1):429. [Google Scholar] | [Crossref] - Yan M, Zhou Y, Cao Y, Li Z, Lu X, Pang B.
Salmonella enterica subsp. II serovar 4,5,12:a:–may cause gastroenteritis in humans. Gut Microbes 2022;14((1)):2089007. [Google Scholar] | [Crossref] - Lai YR, Chang YF, Ma J, Chiu CH, Kuo ML, Lai CH. From DNA damage to cancer progression:effects of cytolethal distending toxin. Front Immunol 2021;12:760451. [Google Scholar] | [Crossref]
- Wang D, Su Y, Gao F, Guo S, Yang J, Wu D. Comparison of ST19
Salmonella Typhimurium and ST34Salmonella I,4,[5],12:i:–:trade-offs between plasmid-mediated resistance and virulence. LWT 2025;225. [Google Scholar] | [Crossref]