


ABSTRACT
Background and Aim: Imerslund–Gräsbeck syndrome (IGS) is a rare autosomal recessive disorder characterized by selective intestinal cobalamin malabsorption in humans and several canine breeds. In Border Collies, IGS is caused by a single cytosine deletion in exon 53 of the
Materials and Methods: A total of 107 clinically healthy Border Collies from private owners and breeding kennels in Thailand were enrolled. Genomic DNA extracted from peripheral blood was genotyped using a newly optimized double-mismatch allele-specific multiplex PCR assay designed to discriminate wild-type and mutant
Results: Of the 107 dogs examined, 106 were homozygous wild-type (WT;
Conclusion: This study provides the first population-level screening data for the
Keywords: Border Collies,
INTRODUCTION
Cobalamin (B12) is an essential water-soluble vitamin involved in multiple physiological processes, including hematopoiesis, neural transmission [1], nucleic acid synthesis, mitochondrial metabolism, intestinal microbial regulation [2], and antioxidant defense [3]. It functions as a cofactor for enzymes responsible for methylmalonic acid degradation and methionine synthesis [4]. In most carnivores, cobalamin is obtained predominantly from animal-derived dietary sources [5]. During digestion, dietary cobalamin initially binds to haptocorrin and is subsequently released by pancreatic enzymes in the intestine, enabling its interaction with the Cubam receptor located on the apical surface of ileal enterocytes. Following cellular uptake, cobalamin binds to transcobalamin, which mediates its transport throughout the systemic circulation [6, 7].
Cubam is a membrane-bound endocytic receptor expressed primarily in the terminal ileum, where it plays a central role in intestinal cobalamin absorption [6]. It is also expressed in the renal proximal tubule and participates in the reabsorption of filtered proteins, particularly albumin [8]. Cubam is a multimeric receptor complex composed of two subunits, amnionless (
In 2013, Fyfe
Despite the established association between
The aim of this study was to determine the genotypic distribution and allele frequency of the
MATERIALS AND METHODS
Ethical approval
The study was conducted in full compliance with national and institutional guidelines for the care and use of research animals. All procedures involving animals were reviewed and approved by the Institutional Animal Care and Use Committee (IACUC), Faculty of Veterinary Medicine, Kasetsart University, Thailand (Approval No. ACKU67-VET-064). The protocol adhered to the ethical principles outlined in the Guide for the Care and Use of Laboratory Animals, the Animal Welfare Act of Thailand, and the Animal Research: Reporting of In vivo Experiments 2.0 guidelines for reporting animal research.
Blood sampling was minimally invasive and performed exclusively by licensed veterinarians. No sedation or anesthesia was required for routine venipuncture; however, appropriate gentle manual restraint was used to minimize stress and ensure animal safety. Each owner (private and kennel owners) provided written informed consent for the collection of blood samples and the use of the animals’ genetic information strictly for research purposes. All samples were handled according to standard biosafety and biosecurity practices. Owner details and kennel identifiers were kept confidential, and no person or identifying information is disclosed in the publication. The IACUC verified that the study posed minimal risk, involved non-terminal and non-invasive sampling, and met all ethical requirements for genetic screening in companion animals.
Study period and location
The research project was conducted from June 2024 to March 2025. All experiments were performed in the molecular research laboratory of the Department of Physiology, Faculty of Veterinary Medicine, Kasetsart University, Thailand (13.8446° N, 100.5773° E). Throughout all laboratory procedures, the room temperature was maintained at 22–25°C, and to ensure consistency, standard laboratory equipment was calibrated regularly. To minimize variability, the timing of each experimental step was strictly followed according to the validated protocol.
Animals and sample collection
Border Collies were selected based on phenotypic characteristics defined in the Fédération Cynologique Internationale breed standard No. 297 to ensure breed authenticity. All dogs had an unknown
A total of 107 dogs, comprising 33 privately owned dogs and 74 dogs from seven breeding kennels, representing both pedigree and non-pedigree backgrounds, were enrolled. Participation was voluntary, and owners and kennel managers provided informed consent for sample collection (Table 1). Pedigree information for dogs originating from registered breeding kennels was verified using breeder- and kennel-provided records, while pedigree data for privately owned dogs were obtained from owner-provided documentation when available.
Table 1. Distribution of dogs by source, sex, and age in the study population.
| Source of dogs | Total (n) | Male, n (%) | Female, n (%) | Age range (months to years) |
|---|---|---|---|---|
| Privately owned dogs | 33 | 18 (54.55) | 15 (45.45) | 3 months to 10 years |
| Kennel A | 49 | 30 (61.22) | 19 (38.78) | 3 months to 7 years |
| Kennel B | 5 | 3 (60) | 2 (40) | 3–7 months |
| Kennel C | 8 | 3 (37.5) | 5 (62.5) | 6 months to 5 years |
| Kennel D | 4 | 0 (0) | 4 (100) | 10 months to 2 years |
| Kennel E | 2 | 2 (100) | 0 (0) | 4–5 years |
| Kennel F | 2 | 0 (0) | 2 (100) | 5–6 months |
| Kennel G | 4 | 1 (25) | 3 (75) | 4 months to 3 years |
| Total | 107 | 57 (53.27) | 50 (46.73) | – |
Peripheral blood samples (1.5 mL per dog) were collected using minimally invasive venipuncture with gentle manual restraint. Samples were collected in tubes containing ethylenediaminetetraacetic acid and stored at 4°C until genomic DNA extraction.
DNA extraction
Genomic DNA extraction was performed under controlled environment conditions (22–25°C) using the QIAamp DNA Blood Mini Kit (Qiagen GmbH, Hilden, Germany) according to the manufacturer’s instructions. The concentration and purity of DNA were evaluated using a NanoDrop™ 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), with a typical concentration of 50–80 ng/µL and acceptable A260/A280 ratios between 1.8 and 2.0.
Samples with concentrations below the minimum concentration threshold and a low A260/A280 ratio were re-extracted and subsequently evaluated by agarose gel electrophoresis to confirm DNA integrity. High-quality DNA samples were aliquoted and stored at −20°C for long-term preservation, and stability was maintained for up to 12–18 months without detectable degradation.
Primer design
This study developed a new set of primers using the integration of double-mismatch allele-specific primers with a multiplex PCR assay. Targeted sequences were identified using the Primer-BLAST tool (NCBI). All primers were designed using sequences deposited in GenBank under accession No. NC_006584.3.
In silico analysis of primer secondary structures, BLAST alignment results, and potential intermolecular interactions, including hairpin loops, self-dimers, and primer-dimer formation, were performed using the OligoAnalyzer™ Tool (Integrated DNA Technologies, Coralville, IA, USA). To ensure adequate purity for downstream PCR applications, oligonucleotides were synthesized and purified using the high-affinity purification method (Biobasic Inc., Markham, ON, Canada) (Table 2).
Table 2. Primer sequences used for
| Primer name | Nucleotide sequence (5′–3′) | Tm (°C) | GC (%) | Product |
|---|---|---|---|---|
| Forward-Internal | AGCTCACTTACAACTCGGAGC | 60.07 | 52.38 | Internal control amplicon (611–612 bp) |
| Reverse-Common | TATGCCATGCCCTTTTGCCTA | 60.06 | 47.62 | — |
| Forward-WT | GGATTTTATGCTACATGGAACACGC | 61.01 | 44 | Wild-type allele amplicon (371 bp) |
| Reverse-Common | TATGCCATGCCCTTTTGCCTA | 60.06 | 47.62 | — |
| Forward-MUT | GGATTTTATGCTACATGGAACACGA | 59.70 | 40 | Mutant allele amplicon (370 bp) |
| Reverse-Common | TATGCCATGCCCTTTTGCCTA | 60.06 | 47.62 | — |
The mismatched bases at the 3′ terminus and penultimate site of allele-specific primers (Forward-WT and Forward-MUT) are shown in bold. WT = wild-type, MUT = mutant, bp = base pair.
A specific primer set consisting of forward wild-type allele-specific (Forward-WT), forward mutant allele-specific (Forward-MUT), and common reverse (Reverse-Common) primers was designed to distinguish target alleles based on a single-nucleotide polymorphism (SNP) region in the
The penultimate position of each allele-specific primer incorporated a second mismatch, where adenine was substituted with guanine. This mismatch reduces heteroduplex stability between the primer and the non-target allele, thereby reducing non-specific amplification [24]. This modification exploits the destabilizing properties of mismatched nucleotides to enhance allele discrimination [25]. The destabilization of primer–template pairs is nucleotide-pair specific, and optimal specificity requires a balance between mismatches at the 3′ terminus and the penultimate site [24].
To achieve high reaction specificity, allele-specific primers were designed based on established

Figure 1. Schematic representation of primer design for single nucleotide polymorphism (SNP) genotyping of the
Schematic representation of primer design for single nucleotide polymorphism (SNP) genotyping of the
To further improve assay reliability, a forward-internal control primer (Forward-Internal) was integrated with the common reverse primer to ensure consistent amplification of an internal control fragment across all samples. Following optimization, the assay enabled co-amplification of a 371-bp wild-type allele fragment, a 370-bp mutant allele fragment, and a 611–612-bp internal control fragment (Figure 1a).
The
The second reaction selectively amplified the mutant allele and contained identical reagents, except Forward-WT was replaced with Forward-MUT (Table 3). Amplification was performed using a MiniAmp Plus Thermal Cycler (Thermo Fisher Scientific, Waltham, MA, USA) with standardized cycling conditions, consisting of an initial denaturation at 95C for 5 min, followed by 35 cycles of denaturation at 95°C for 30 s, primer annealing at 59°C for 2 min, and extension at 72°C for 1 min, with a final extension step at 72°C for 10 min.
Table 3. Composition of a 25 L PCR reaction mixture for
| Reagent | Stock concentration | Volume added (µL) | Final concentration in 25 µL |
|---|---|---|---|
| Platinum® Taq PCR buffer | 10 × | 2.5 | 1× |
| dNTP mixture | 2 mM | 2.5 | 0.2 mM each |
| MgCl2 | 50 mM | 0.75 | 1.5 mM |
| Forward-Internal primer | 10 µM | 0.75 | 0.3 µM |
| Forward-WT primer | 10 µM | 0.75 | 0.3 µM |
| Forward-MUT primer | 10 µM | 0.75 | 0.3 µM |
| Reverse-Common primer | 10 µM | 0.75 | 0.3 µM |
| Platinum® Taq DNA polymerase | 5 U/µL | 0.1 | 0.5 U |
| Template DNA | 50–80 ng/µL | 1.0 | 50–80 ng |
| Nuclease-free water | – | 15.9 | – |
| Total | 25.0 |
* Forward-WT and Forward-MUT primers were added to a separate reaction tube.
PCR products were resolved on 1.5% agarose gels prepared in 1× TBE buffer and pre-stained with ethidium bromide (0.5 µg/mL). Electrophoresis was conducted at 100 V for approximately 30 min, and fragments were visualized under UV illumination using a Gel Doc™ EZ Documentation System (Bio-Rad Laboratories, Hercules, CA, USA).
Synthetic mutated DNA fragments of CUBN
Due to the absence of naturally occurring homozygous mutant individuals, a synthetic 924-bp DNA fragment containing the

Figure 2. Schematic of the synthetic 924-bp DNA fragment containing the
Schematic of the synthetic 924-bp DNA fragment containing the
Sequencing
Following PCR-based
Consensus sequences were generated using SnapGene® Viewer (GSL Biotech LLC, San Diego, CA, USA) and aligned with the reference
Statistical analysis
Prevalence and allele frequencies were calculated as proportions, and 95% confidence intervals (CI) were estimated using the Wilson score method to improve accuracy for rare events in moderate sample sizes.
RESULTS
Technical validation of combined double-mismatch allele-specific and multiplex PCR assays
All 107 samples were successfully and reliably genotyped for the
The assay performance was further validated using synthetic mutated DNA fragments. As expected, this positive control generated a
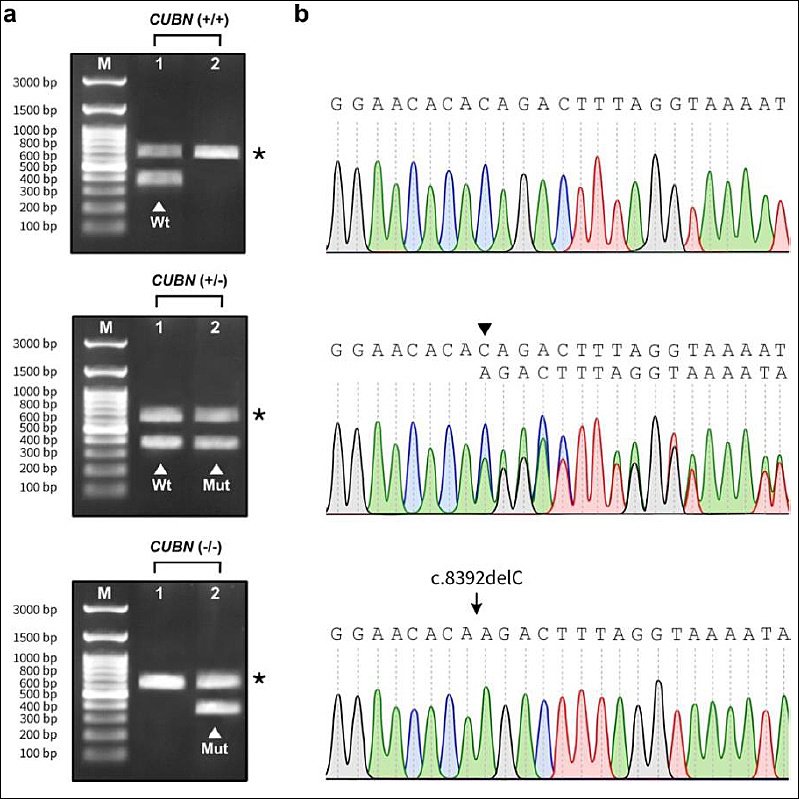
Figure 3. Validation of
Validation of
The multiplex double-mismatch allele-specific PCR assay demonstrated high reproducibility across all replicates, with 100% concordance observed. Genotyping results for all 107 samples were fully consistent with those obtained by Sanger sequencing, with no false-positive or false-negative findings detected (Figure 3b). Collectively, these results confirm that the proposed assay is highly efficient, sensitive, and specific, providing a reliable detection platform for the targeted
CUBN genotype profiles associated with IGS
The genotypic distribution and allele frequencies of the
Table 4. Distribution of
| Category | Genotype/Allele | Frequency (%) | Wilson 95% CI |
|---|---|---|---|
| Genotype frequencies | 99.1 | 94.8–99.8 | |
| 0.9 | 0.16–5.0 | ||
| 0 | 0–4.3 | ||
| Allelic frequencies | Wild-type allele | 99.6 | 98.2–99.9 |
| Mutant allele | 0.4 | 0.07–1.8 |
The calculated allele frequencies indicated a predominance of the wild-type allele at 99.6% (95% CI: 98.2%–99.9%), whereas the mutant allele frequency was estimated at 0.4% (95% CI: 0.07%–1.8%). Taken together, the absence of homozygous mutant individuals and the very low mutant allele frequency suggest that the estimated prevalence of IGS is relatively low in this Border Collie population.
DISCUSSION
Development and validation of the genotyping approach
This study aimed to investigate
A forward-internal control primer was designed to target a conserved region within intron 52 without nucleotide mismatches. The inclusion of parallel internal control amplification provided an additional reliability feature that is not commonly incorporated in previously published allele-specific PCR designs for IGS. When paired with the common reverse primer, this internal control primer consistently amplified a fragment of 611- or 612-bp across the SNP region in all samples. The internal control fragment was clearly visualized and readily distinguished from the smaller allele-specific products (370-bp for the mutant allele and 371-bp for the WT allele) by agarose gel electrophoresis (Figure 3a).
To further validate assay performance, a custom-designed 924-bp synthetic DNA fragment representing the mutant allele was included to confirm the detection of homozygous mutant genotypes. This strategy represents a novel control approach that enables reliable interpretation of results in populations in which homozygous mutants are extremely rare. This is the first study to adapt and validate a double-mismatch allele-specific PCR assay for the rapid and cost-efficient detection of
Practical implications for genetic screening and disease control
The findings of this study have important practical implications for genetic counselors, breeders, and clinicians. Integration of these results into genetic counseling frameworks allows collaboration with breeders to develop long-term preventive breeding strategies that maintain disease-free populations while preserving genetic diversity. Routine genetic screening before breeding can help avoid carrier-to-carrier matings and prevent the production of affected offspring. In addition, this diagnostic approach provides clinicians with a practical tool for incorporating genetic testing into routine diagnostic protocols for dogs presenting compatible clinical signs or originating from high-risk lineages.
Population structure and CUBN genotype distribution
A genetic survey was conducted on 107 Border Collies in Thailand, comprising 33 privately owned dogs with unrelated parental origins and 74 dogs from seven breeding kennels. Kennel A contributed the largest proportion of dogs (n = 49), with approximately one-third sharing common parentage, whereas kennels B (n = 5), C (n = 8), D (n = 4), E (n = 2), F (n = 2), and G (n = 4) contributed smaller numbers. Pedigree verification demonstrated that the study population represented a heterogeneous genetic pool. Nearly half of the dogs were direct descendants of Border Collies originating from the United States, while the remaining dogs traced ancestry to Australia, Russia, Austria, France, Hungary, Italy, and the United Kingdom.
Genotypic analysis revealed a predominance of the WT
Comparison with international studies and population genetics considerations
A cohort study from Switzerland involving 203 Border Collies reported a substantially higher carrier frequency of 6.2% for the
Analysis of imported and locally bred Border Collies in Thailand underscores the influence of international breeding lines on the distribution of hereditary disorders in emerging populations. The relatively low carrier frequency observed in this cohort is likely attributable to a geographically diverse population structure composed of dogs imported from multiple countries and their descendants. Such diversity establishes a heterogeneous founder population, mitigating founder effects and reducing
Study limitations and future perspectives
Despite the predominance of the WT
Second, the use of convenience sampling rather than randomized selection may have resulted in uneven representation of specific kennels, introducing potential selection bias and limiting generalizability to the broader Thai Border Collie population. Third, pedigree analysis was not performed because comprehensive multigenerational lineage records were unavailable for many dogs, particularly privately owned and adopted individuals. Consequently, future studies incorporating larger sample sizes, well-documented pedigrees, and stratified random sampling are required to refine prevalence estimates and better characterize transgenerational inheritance patterns of IGS.
Integration with existing genetic screening data
This study complements recent genetic screening efforts targeting multidrug resistance 1 (
CONCLUSION
This study successfully identified
The validated assay provides a rapid, reliable, and cost-efficient diagnostic tool for routine
Several limitations should be acknowledged. The moderate sample size may limit the detection of rare genotypes and reduce statistical power. Convenience sampling and uneven representation among kennels may introduce selection bias, restricting extrapolation to the entire Thai Border Collie population. Furthermore, incomplete pedigree information for many dogs prevented detailed transgenerational inheritance analysis.
Future studies should incorporate larger, randomly sampled cohorts with comprehensive pedigree data to refine prevalence estimates and better characterize population-level inheritance patterns of IGS. Integration of this PCR-based approach with high-throughput genotyping platforms may further enhance large-scale surveillance. Longitudinal studies combining genetic data with clinical and biochemical assessments would also improve understanding of genotype–phenotype relationships.
Overall, this study provides the first population-level insight into
DATA AVAILABILITY
Additional data supporting the findings of this study are available from the corresponding author upon reasonable request.
AUTHORS’ CONTRIBUTIONS
CL: Examined the clinical condition of the animals, collected blood samples, designed the study, performed the experiments, analyzed data, drafted, reviewed, and edited the manuscript. NA: designed the study, analyzed data, reviewed and edited the manuscript. NP: Provided technical laboratory support, reviewed and edited the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors would like to thank all dog owners and breeders involved in the study. This research was financially supported by the Faculty of Veterinary Medicine, Kasetsart University, Thailand (Grant no. VET.KU2024-RPDF03).
REFERENCES
- Wolffenbuttel BH, McCaddon A, Ahmadi KR, Green R. A brief overview of the diagnosis and treatment of cobalamin (B12) deficiency. Food Nutr Bull 2024;45((1 Suppl)):S40-S59. [Google Scholar] | [Crossref]
- Hossain KS, Amarasena S, Mayengbam S. B vitamins and their roles in gut health. Microorganisms 2022;10(6):1168. [Google Scholar] | [Crossref]
- Offringa AK, Bourgonje AR, Schrier MS, Deth RC, van Goor H. Clinical implications of vitamin B12 as redox-active cofactor. Trends Mol Med 2021;27((10)):931-934. [Google Scholar] | [Crossref]
- Nexo E, Parkner T. Vitamin B12-related biomarkers. Food Nutr Bull 2024;45((1 Suppl)):S28-S33. [Google Scholar] | [Crossref]
- Moravcová M, Siatka T, Krčmová LK, Matoušová K, Mladěnka P. Biological properties of vitamin B12. Nutr Res Rev 2025;38(1):338-370. [Google Scholar] | [Crossref]
- Kather S, Grützner N, Kook PH, Dengler F, Heilmann RM. Review of cobalamin status and disorders of cobalamin metabolism in dogs. J Vet Intern Med 2020;34((1)):13-28. [Google Scholar] | [Crossref]
- Kozyraki R, Verroust P, Cases O. Cubilin, the intrinsic factor-vitamin B12 receptor. Vitam Horm 2022;119:65-119. [Google Scholar] | [Crossref]
- Gburek J, Konopska B, Gołąb K. Renal handling of albumin—from early findings to current concepts. Int J Mol Sci 2021;22(11):5809. [Google Scholar] | [Crossref]
- Aminoff M, Carter JE, Chadwick RB, Johnson C, Gräsbeck R, Abdelaal MA, Broch H, Jenner LB, Verroust PJ, Moestrup SK, de la Chapelle A, Krahe R. Mutations in
CUBN , encoding the intrinsic factor-vitamin B12 receptor, cubilin, cause hereditary megaloblastic anaemia 1. Nat Genet 1999;21(3):309-313. [Google Scholar] | [Crossref] - Tanner SM, Aminoff M, Wright FA, Liyanarachchi S, Kuronen M, Saarinen A, Massika O, Mandel H, Broch H, de la Chapelle A. Amnionless, essential for mouse gastrulation, is mutated in recessive hereditary megaloblastic anemia. Nat Genet 2003;33(3):426-429. [Google Scholar] | [Crossref]
- He Q, Madsen M, Kilkenney A, Gregory B, Christensen EI, Vorum H, Højrup P, Schäffer AA, Kirkness EF, Tanner SM, de la Chapelle A, Giger U, Moestrup SK, Fyfe JC. Amnionless function is required for cubilin brush-border expression and intrinsic factor-cobalamin (vitamin B12) absorption in vivo. Blood 2005;106(4):1447-1453. [Google Scholar] | [Crossref]
- Gold AJ, Scott MA, Fyfe JC. Failure to thrive and life-threatening complications due to inherited selective cobalamin malabsorption effectively managed in a juvenile Australian shepherd dog. Can Vet J 2015;56(10):1029-1034. [Google Scholar] | [Crossref]
- Fyfe JC, Giger U, Hall CA, Jezyk PF, Klumpp SA, Levine JS, Patterson DF. Inherited selective intestinal cobalamin malabsorption and cobalamin deficiency in dogs. Pediatr Res 1991;29((1)):24-31. [Google Scholar] | [Crossref]
- Fordyce HH, Callan MB, Giger U. Persistent cobalamin deficiency causing failure to thrive in a juvenile beagle. J Small Anim Pract 2000;41(9):407-410. [Google Scholar] | [Crossref]
- Kook PH, Drögemüller M, Leeb T, Howard J, Ruetten M. Degenerative liver disease in young beagles with hereditary cobalamin malabsorption because of a mutation in the
cubilin gene. J Vet Intern Med 2014;28((2)):666-671. [Google Scholar] | [Crossref] - Fyfe JC, Hemker SL, Venta PJ, Stebbing B, Giger U. Selective intestinal cobalamin malabsorption with proteinuria (Imerslund-Gräsbeck syndrome) in juvenile beagles. J Vet Intern Med 2014;28((2)):356-362. [Google Scholar] | [Crossref]
- Murtagh K, Batchelor D, German A, Piviani M, Silvestrini P. Congenital cobalamin malabsorption (Imerslund-Gräsbeck syndrome) in two beagles in the UK. Vet Rec Case Rep 2015;3(1):e000201. [Google Scholar] | [Crossref]
- Battersby IA, Giger U, Hall EJ. Hyperammonaemic encephalopathy secondary to selective cobalamin deficiency in a juvenile Border collie. J Small Anim Pract 2005;46(7):339-344. [Google Scholar] | [Crossref]
- Fyfe JC, Hemker SL, Venta PJ, Fitzgerald CA, Outerbridge CA, Myers SL, Giger U. An exon 53 frameshift mutation in
CUBN abrogates cubam function and causes Imerslund-Gräsbeck syndrome in dogs. Mol Genet Metab 2013;109(4):390-396. [Google Scholar] | [Crossref] - Owczarek-Lipska M, Jagannathan V, Drögemüller C, Lutz S, Glanemann B, Leeb T, Kook PH. A frameshift mutation in the cubilin gene (
CUBN ) in Border collies with Imerslund-Gräsbeck syndrome (selective cobalamin malabsorption). PLoS One 2013;8(4):e61144. [Google Scholar] | [Crossref] - Lutz S, Sewell AC, Reusch CE, Kook PH. Clinical and laboratory findings in Border collies with presumed hereditary juvenile cobalamin deficiency. J Am Anim Hosp Assoc 2013;49(3):197-203. [Google Scholar] | [Crossref]
- Drögemüller M, Jagannathan V, Howard J, Bruggmann R, Drögemüller C, Ruetten M, Leeb T, Kook PH. A frameshift mutation in the cubilin gene (
CUBN ) in beagles with Imerslund-Gräsbeck syndrome (selective cobalamin malabsorption). Anim Genet 2014;45(1):148-150. [Google Scholar] | [Crossref] - Mizukami K, Yabuki A, Kohyama M, Kushida K, Rahman MM, Uddin MM, Sawa M, Yamato O. Molecular prevalence of multiple genetic disorders in Border collies in Japan and recommendations for genetic counselling. Vet J 2016;214:21-23. [Google Scholar] | [Crossref]
- Bui M, Liu Z. Simple allele-discriminating PCR for cost-effective and rapid genotyping and mapping. Plant Methods 2009;5:1. [Google Scholar] | [Crossref]
- Huang K, Zhang J, Li J, Qiu H, Wei L, Yan Y, Wang C. Exploring the impact of primer-template mismatches on PCR performance of DNA polymerase varying in proofreading activity. Genes 2024;15((2)):215. [Google Scholar] | [Crossref]
- Lerdkrai C, Phungphosop N. Prevalence of the MDR1 gene mutation in herding dog breeds and Thai Ridgebacks in Thailand. Vet World 2021;14(11):3015-3020. [Google Scholar] | [Crossref]
- Lerdkrai C, Phungphosop N. A novel multiplex polymerase chain reaction assay for the genotypic survey of the non-homologous end-joining factor 1 associated with Collie eye anomaly in Thailand. Vet World 2022;15(1):132-139. [Google Scholar] | [Crossref]
- Lerdkrai C, Phungphosop N. Multiplex allele-specific polymerase chain reaction-based prevalence study of canine trapped neutrophil syndrome in Thailand. Vet World 2023;16(11):2271-2276. [Google Scholar] | [Crossref]