


ABSTRACT
Background and Aim: Essential oils (EOs) are promising natural modifiers of rumen fermentation and methane production; however, their volatility and rapid degradation limit their effectiveness. Microencapsulation can shield bioactive compounds and allow controlled release. Insect-derived proteins, especially from black soldier fly (BSF;
Materials and Methods: A completely randomized design was used with five dietary treatments containing M-LEO at 0, 2, 4, 6, and 8% of total dry matter (DM) substrate.
Results: M-LEO showed high encapsulation efficiency (85.2%) and significant bioactive content. Supplementing with M-LEO notably improved gas production kinetics and nutrient degradability, with optimal effects at 6% of total DM. At this level, IVDMD and IVOMD increased by up to 11.5% and 10.5%, respectively. Total VFA and propionate concentrations rose significantly (p < 0.05), while acetate proportion and the acetate-to-propionate ratio decreased. Rumen pH and NH3-N levels stayed within optimal ranges and were unaffected by treatment. Methane production was substantially reduced, with decreases of up to 48.8% at 48 h compared to the control. Additionally, M-LEO boosted populations of key cellulolytic bacteria (
Conclusion: Microencapsulation of lemongrass oil with BSF protein effectively enhances rumen fermentation efficiency and significantly decreases methane emissions
Keywords: black soldier fly protein, climate-smart ruminant nutrition, essential oil microencapsulation, lemongrass oil, methane mitigation, rumen fermentation, sustainable feed additive,
INTRODUCTION
Currently, a wide variety of plant-derived bioactive compounds (BCs), including phenolics, flavonoids, and especially essential oils (EOs), have been shown to influence rumen fermentation processes. As a result, plant-based extracts have gained significant interest as natural feed additives for ruminants [1]. EOs are complex mixtures of volatile, lipophilic secondary metabolites produced by plants and are responsible for their characteristic aroma and color [2]. These compounds are typically extracted using hydro-distillation, solvent extraction, or similar methods [3]. Due to their antimicrobial properties, EOs are increasingly recognized as natural alternatives to synthetic antibiotics in animal nutrition [4]. Furthermore, several EOs demonstrate strong antioxidant activity; for instance, EOs from clove, oregano, parsley, and tarragon have been shown to inhibit 2,2-diphenyl-1-picrylhydrazyl (DPPH) radicals by up to 50% [5]. Collectively, these properties enable EOs to affect key rumen fermentation traits that are vital for ruminant productivity and health [6].
Lemongrass (
Microencapsulation involves trapping small particles or droplets within a homogeneous or heterogeneous matrix or coating material to form microcapsules [13]. This technology enhances the handling, stability, and delivery efficiency of BCs [14]. The encapsulated core material is gradually released through the capsule wall, enabling sustained and controlled delivery. A wide variety of wall materials can be used, including natural polymers (e.g., cellulose, chitosan, and starch) and synthetic polymers (e.g., polyethylene, polyester, and nylon), depending on the desired properties of the final product [15]. Recently, insects have emerged as a promising alternative source of wall materials due to their high-quality protein content [16, 17]. In this study, proteins derived from the black soldier fly (BSF;
Plant-derived BC, especially EO, has been thoroughly studied for its ability to influence rumen fermentation and reduce enteric methane emissions. Many
Meanwhile, insect-derived proteins, especially those from BSF larvae, have gained recognition as sustainable feed ingredients due to their high protein content, favorable amino acid profile, and their role in circular bioeconomy systems. Despite these benefits, the potential of BSF protein as a functional wall material for microencapsulation of EO has not been thoroughly investigated. Currently, there is a noticeable lack of information on how insect-protein–based encapsulation systems affect the release of essential oil bioactive compounds in the rumen environment, their interactions with rumen microbial communities, and their subsequent impact on fermentation efficiency and methane production. Additionally, limited data exist on the best inclusion levels of microencapsulated EO using novel biopolymer walls that enhance fermentation benefits while reducing negative effects on rumen function. This knowledge gap hinders the development of scalable, climate-smart, and biologically efficient feed additives aimed at methane mitigation in ruminant production systems.
To address these gaps, this study aimed to evaluate the potential of BSF protein–based microencapsulation as a new delivery system for lemongrass essential oil in ruminant nutrition. Specifically, it sought to examine the effects of microencapsulated-lemongrass oil (M-LEO) on
MATERIALS AND METHODS
Ethical approval
All experimental procedures involving animals were reviewed and approved by the Institutional Animal Care and Use Committee of Khon Kaen University (IACUC-KKU), Thailand (approval number: IACUC-KKU-86/66), and the Institute of Animals for Scientific Purpose Development (IAD), Thailand (approval number: U1-07387-2561). Animal handling, rumen fluid collection, and
Rumen fluid was collected non-invasively from healthy dairy cattle maintained under standard management practices, causing minimal stress and discomfort to the animals. All experimental protocols adhered to the ethical principles outlined by the Animal Research: Reporting of
Study period and location
This research was carried out from October 2024 to February 2025 at the Tropical Feed Resources Research and Development Center, Department of Animal Science, Faculty of Agriculture, Khon Kaen University, Khon Kaen, Thailand.
Preparation of M-LEO
The microencapsulation process followed the method described by Phupaboon
Diet formulation and chemical composition analysis
The basal diet consisted of concentrate and roughage sources, as shown in Table 1. The concentrate was formulated with cassava chips, rice bran meal, palm kernel meal, soybean meal, urea, sulfur, mineral premix, and salt, providing a crude protein content of 14.6% on a dry matter (DM) basis. Rice straw was used as the roughage source, containing 2.4% crude protein, 78.9% neutral-detergent fiber (NDF), and 52.6% acid-detergent fiber (ADF).
Table 1. Dietary ingredients and nutritional composition.
| Items | Concentrate | Rice straw | Black soldier fly extract | M-LEO |
|---|---|---|---|---|
| Ingredients (% as fed) | - | - | - | - |
| Cassava chip | 54 | - | - | - |
| Rice bran meal | 17 | - | - | - |
| Palm kernel meal | 13 | - | - | - |
| Soybean meal | 10.5 | - | - | - |
| Urea | 2.5 | - | - | - |
| Sulphur | 1 | - | - | - |
| Mineral premixed | 1 | - | - | - |
| Salt | 1 | - | - | - |
| Chemical compositions | - | - | - | - |
| Dry matter (DM, %) | 90.5 | 89.4 | 95.1 | 92.2 |
| Organic matter | 92.2 | 85.4 | 93.5 | 93.7 |
| Crude protein | 14.6 | 2.4 | 50 | 21.9 |
| Neutral-detergent fiber | 20.4 | 78.9 | 53.1 | 30.2 |
| Acid-detergent fiber | 12.6 | 52.6 | 38.9 | 23.5 |
| Bioactive compounds | - | - | - | - |
| TPC (mg GAE/g DM) | - | - | - | 1927.4 |
| TFC (mg QUE/g DM) | - | - | - | 789.2 |
| Antioxidant capacities | - | - | - | - |
| ABTS (%) | - | - | - | 12.8 |
| DPPH (%) | - | - | - | 78.1 |
| FRAP (g TROE/g DM) | - | - | - | 20.6 |
| Encapsulation efficiency (%) | - | - | - | 85.2 |
M-LEO = Microencapsulated-lemongrass oil, TPC = Total phenolic compounds, TFC = Total flavonoid compounds, ABTS = 2’-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid), DPPH = 2, 2-diphenyl-1-picrylhydrazyl, FRAP = Ferric reducing antioxidant power.
1 Mineral premixed (contains per kg): vitamin A, 10,000,000 IU; vitamin D, 1,600,000 IU; vitamin E, 70,000 IU; Fe, 50 g; Mn, 40 g; Zn, 40 g; Cu, 10 g; I, 0.5 g; Se, 0.1 g; Co, 0.1 g.
Chemical analyses of concentrate, rice straw, and M-LEO were performed following AOAC methods [20] for DM (method no. 967.03), organic matter (OM; method no. 942.05), and crude protein (CP; method no. 984.13). Fiber fractions (NDF and ADF) were determined using an Ankom fiber analyzer (Ankom Technology Co., NY, USA) following the protocol of Van Soest
Determination of BC and antioxidant capacity
M-LEO was analyzed for BC, including total phenolic and flavonoid contents, as well as antioxidant capacities. Total phenolic content was determined using the modified Folin–Ciocalteu method with gallic acid as the standard [22]. Total flavonoid content was quantified using a calibration curve generated from quercetin standards prepared by two-fold serial dilutions in ethanol [23]. Antioxidant activity was assessed using the DPPH radical scavenging assay [24], ABTS radical scavenging assay [25], and ferric reducing antioxidant power (FRAP) assay [26].
Encapsulation efficiency (EE) was determined using the equation: EE (%) = 100 × (total phenolic compounds extracted/total phenolic compounds encapsulated), as reported by Phupaboon
In vitro experimental design and treatments
A completely randomized design was used with five dietary treatments, each involving M-LEO supplementation at 0, 2, 4, 6, and 8% of total DM substrate. Each treatment had three replicates and was tested at two incubation times, along with three blank replicates for each measured parameter.
Animals, rumen fluid collection, and inoculum preparation
Rumen inoculum was collected from four Holstein-crossbred dairy cattle (Holstein Friesian × Thai native) with an average body weight of 500 ± 20 kg. The animals were fed rice straw
In vitro rumen incubation procedure
The
Measurement of gas production kinetics
Gas production was measured using a 20 mL glass precision syringe at 1, 2, 4, 6, 8, 12, 24, 48, 72, and 96 h of incubation. Gas kinetic parameters were estimated with the nonlinear regression model of Ørskov and McDonald [29]:
Y = a + b (1 − e-ct),
where Y represents gas production (mL) at time t, a is gas production from the soluble fraction, b is gas production from the insoluble fraction, c is the fractional gas production rate, and t is incubation time (h).
Rumen fermentation characteristics and methane analysis
Immediately after bottle opening at 12, 24, and 48 h, rumen pH was measured using a calibrated digital pH meter (HANNA Instruments HI 8424, Singapore). VFAs and ammonia-nitrogen (NH2-N) concentrations were determined after centrifuging filtered rumen fluid samples at 16,000 ×
NH2-N concentration was measured spectrophotometrically using a LabAssay™ Ammonia kit (FUJIFILM Wako Pure Chemical Corp., Japan) with absorbance read at 630 nm. Methane production was analyzed with gas chromatography (GC-17A System, Shimadzu Co. Ltd., Kyoto, Japan) equipped with a thermal conductivity detector, following the method described by Pattra
In vitro nutrient degradability
Microbial DNA extraction and quantitative polymerase chain reaction (PCR) analysis
Total genomic DNA was extracted from about 1 mL of rumen fluid using the QIAamp Fast DNA Stool Mini Kit (Qiagen, Hilden, Germany). DNA purity and concentration were measured with a Nanodrop spectrophotometer (Thermo Scientific, Waltham, MA, USA). Quantitative real-time PCR was conducted using Luna® Universal qPCR Master Mix with species-specific primers targeting key rumen bacteria and methanogens, including R
Statistical analysis
All data were analyzed using the general linear model procedure in SAS software (version 9.4; SAS Institute, Cary, NC, USA) under a completely randomized design. The statistical model was Yij = μ + Mi + εij, where μ represents the overall mean, Mi denotes the effect of M-LEO supplementation level, and εij is the residual error. Treatment means were compared using Tukey’s test, and orthogonal polynomial contrasts (linear, quadratic, and cubic) were applied to assess responses to increasing M-LEO levels. Statistical significance was set at p < 0.05 and p < 0.01.
RESULTS
Nutritive composition and EE of M-LEO
M-LEO showed a DM content of 92.2%, with crude protein, NDF, and ADF contents of 21.9%, 30.2%, and 23.5%, respectively. The bioactive profile of M-LEO revealed high levels of total phenolic compounds (1927.4 mg GAE/g DM) and total flavonoid compounds (789.2 mg QUE/g DM). The EE, based on the recovery of total phenolic compounds within the encapsulated matrix, was 85.2%, demonstrating effective entrapment of bioactive substances (Table 1).
Effects of M-LEO on gas kinetics and nutrient degradability
Supplementation with M-LEO significantly affected cumulative gas production (p < 0.01), as shown in Figure 1. Higher levels of M-LEO led to increased gas production from the immediately soluble fraction (a; p < 0.05), the insoluble fraction (b; p < 0.01), the gas production rate constant (c; p < 0.01), the potential extent of gas production (a + b; p < 0.01), and cumulative gas volume at 96 h (p < 0.01) compared to the control. Additionally, including M-LEO at 2–6% of the total DM substrate improved IVDMD and IVOMD at 12, 24, and 48 h, whereas a reduction was observed at the 8% inclusion level. The most significant improvements in degradability were seen at 48 h with 6% M-LEO supplementation, where IVDMD and IVOMD increased by 11.5% and 10.5%, respectively (Table 2).
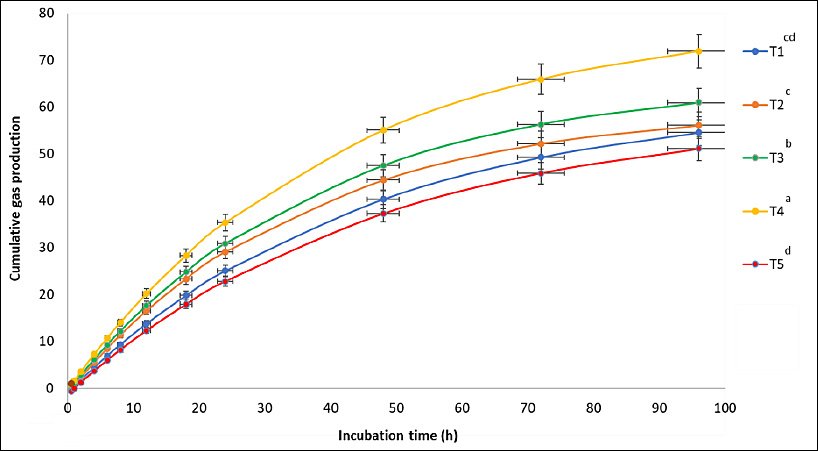
Figure 1. Cumulative gas production curves for the treatment substrates in an
Table 2.
| Treatments | M-LEO | Gas kinetics | Cumulative | IVDMD (% DM) | IVOMD (% DM) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
| |||||||||
|
|
|
| gas | 12 h | 24 h | 48 h | 12 h | 24 h | 48 h | |||
| 1 | 0 | -0.9bc | 62.9bc | 0.022b | 62.0bc | 55.6cd | 51.0b | 55.9bc | 60.9c | 57.9b | 63.0bc | 68.2c |
| 2 | 2 | -1.1cd | 61.2bc | 0.028a | 60.1bc | 58.1c | 51.9b | 57.9ab | 64.0b | 58.9b | 65.0ab | 71.2b |
| 3 | 4 | -0.6b | 66.7b | 0.027a | 66.1b | 62.8b | 53.3ab | 58.3ab | 65.6b | 60.1ab | 65.4ab | 72.8b |
| 4 | 6 | -0.3a | 79.6a | 0.025ab | 79.3a | 73.6a | 56.6a | 59.7a | 68.8a | 63.4a | 67.0a | 76.2a |
| 5 | 8 | -1.3d | 60.0c | 0.021b | 58.8c | 52.5d | 50.6b | 54.6c | 59.5c | 57.1b | 61.7c | 66.8c |
| SEM | 0.1 | 0.45 | 0.01 | 0.46 | 0.34 | 0.32 | 0.26 | 0.24 | 0.33 | 0.27 | 0.23 | |
| Orthogonal polynomials | ||||||||||||
| Linear | <0.01 | <0.01 | 0.27 | <0.01 | <0.01 | 0.01 | 0.01 | <0.01 | 0.01 | 0.01 | <0.01 | |
| Quadratic | 0.02 | <0.01 | <0.01 | <0.01 | <0.01 | 0.33 | 0.7 | 0.91 | 0.35 | 0.79 | 0.74 | |
| Cubic | 0.08 | 0.97 | 0.27 | 0.94 | 0.42 | 0.8 | 0.49 | 0.31 | 0.75 | 0.46 | 0.24 | |
M-LEO = Microencapsulated-lemongrass oil (% of total DM substrate), IVDMD =
1 Gas production kinetics, a = Gas production from the immediately soluble fraction (mL), b = Gas production from the insoluble fraction (mL), c = Gas production rate constant for the insoluble fraction (mL/h), a + b = Potential extent of gas production (mL).
2 Cumulative gasses at 96 h (mL per 0.5 g DM substrate).a–d Means within the same column with different letters are significantly different at p < 0.05.
Methane production
A significant reduction in methane production was observed following M-LEO supplementation at 12 h (p < 0.05; linear effect), 24 h (p < 0.05; quadratic effect), and 48 h (p < 0.01; linear effect), with the most notable effects occurring at supplementation levels of 6%–8% of total DM substrate. Compared to the control group, methane emissions decreased by 27.8%, 58.7%, and 51.2% at 12, 24, and 48 h, respectively, when M-LEO was included at 8% of total DM substrate (Table 3).
Table 3.
| Treatments | M-LEO | VFA (mol/100 mL) | C2:C3 | Total VFA (mmol/L) | Methane production (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
|
|
| ||||||||
| C2 | C3 | C4 | 12 h | 24 h | 48 h | ||||
| 1 | 0 | 67.6a | 22.0b | 10.5 | 3.1a | 103.8b | 7.2a | 6.3a | 4.3a |
| 2 | 2 | 66.5ab | 23.0ab | 10.6 | 2.9ab | 108.9ab | 6.9ab | 6.0ab | 3.6b |
| 3 | 4 | 66.0ab | 23.0ab | 11.0 | 2.9ab | 110.0ab | 6.6ab | 5.2b | 2.8c |
| 4 | 6 | 65.1b | 24.8a | 10.1 | 2.6b | 115.8a | 5.5ab | 3.6c | 2.2d |
| 5 | 8 | 67.0ab | 22.0b | 11.0 | 3.05a | 105.5b | 5.2b | 2.6d | 2.1d |
| SEM | 0.23 | 0.24 | 0.22 | 0.10 | 0.43 | 0.20 | 0.15 | 0.10 | |
| Orthogonal polynomials | |||||||||
| Linear | 0.02 | 0.03 | 0.77 | 0.02 | 0.01 | 0.04 | <0.01 | <0.01 | |
| Quadratic | 0.80 | 0.53 | 0.38 | 0.82 | 0.87 | 0.40 | 0.01 | 1.00 | |
| Cubic | 0.72 | 0.39 | 0.57 | 0.49 | 0.38 | 0.73 | 0.48 | 0.72 | |
M-LEO = Microencapsulated-lemongrass oil (% of total dry matter substrate), VFA = Volatile fatty acids, C2 = Acetate, C3 = Propionate, C4 = Butyrate, C2:C3 = Acetate-to-propionate ratio, SEM = Standard error of the mean. a–c Means within the same column with different letters are significantly different at p < 0.05.
Table 4.
| Treatments | M-LEO | pH | Ammonia-nitrogen (mg/dL) | ||||
|---|---|---|---|---|---|---|---|
|
|
| ||||||
| 12 h | 24 h | 48 h | 12 h | 24 h | 48 h | ||
| 1 | 0 | 6.94 | 6.90 | 6.88 | 16.0 | 16.8 | 18.2 |
| 2 | 2 | 6.93 | 6.94 | 6.92 | 16.8 | 17.6 | 18.4 |
| 3 | 4 | 6.93 | 6.91 | 6.89 | 16.9 | 17.7 | 18.5 |
| 4 | 6 | 6.91 | 6.89 | 6.87 | 16.9 | 17.7 | 18.5 |
| 5 | 8 | 6.92 | 6.90 | 6.88 | 16.8 | 17.8 | 18.6 |
| SEM | 0.03 | 0.04 | 0.04 | 0.15 | 0.16 | 0.12 | |
| Orthogonal polynomials | |||||||
| Linear | 0.20 | 0.57 | 0.58 | 0.05 | 0.19 | 0.24 | |
| Quadratic | 0.59 | 0.23 | 0.20 | 0.13 | 0.36 | 0.46 | |
| Cubic | 0.47 | 0.51 | 0.63 | 0.61 | 0.73 | 0.84 | |
M-LEO = Microencapsulated-lemongrass oil (% of total dry matter substrate), SEM = Standard error of the mean.
Modulation of rumen microbial populations
M-LEO supplementation positively affected rumen microbial dynamics by increasing the abundance of important cellulolytic bacteria, including
Table 5.
| Species (log copies/mL) | IT (h) | M-LEO | SEM | Orthogonal polynomials | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
|
| |||||||||
| 0 | 2 | 4 | 6 | 8 | L | Q | C | |||
|
| 12 | 8.5 | 8.5 | 8.7 | 8.8 | 8.7 | 0.10 | 0.08 | 0.70 | 0.86 |
| 24 | 8.4ab | 8.8ab | 8.3b | 8.9a | 8.6ab | 0.11 | 0.17 | 0.73 | 0.02 | |
| 48 | 8.6b | 9.9ab | 11.0ab | 11.4a | 9.3ab | 0.24 | 0.02 | 0.52 | 0.87 | |
|
| 12 | 9.2 | 9.5 | 9.3 | 9.7 | 9.4 | 0.13 | 0.19 | 0.79 | 0.31 |
| 24 | 8.7b | 9.1ab | 9.1ab | 9.5a | 9.5a | 0.11 | 0.03 | 0.90 | 0.37 | |
| 48 | 9.1b | 9.3ab | 9.4ab | 9.8a | 9.3ab | 0.13 | 0.03 | 0.62 | 0.74 | |
|
| 12 | 8.8 | 8.8 | 9.0 | 9.2 | 8.3 | 0.34 | 0.37 | 0.88 | 0.89 |
| 24 | 7.9 | 9.0 | 9.4 | 9.5 | 7.6 | 0.21 | 0.06 | 0.38 | 0.90 | |
| 48 | 7.8b | 8.9ab | 9.7a | 10.1a | 8.7ab | 0.22 | 0.01 | 0.42 | 0.95 | |
|
| 12 | 8.6 | 9.1 | 9.1 | 9.4 | 8.9 | 0.14 | 0.05 | 0.74 | 0.54 |
| 24 | 8.7c | 9.0bc | 9.3ab | 9.5a | 8.9c | 0.09 | <0.01 | 0.81 | 0.75 | |
| 48 | 9.1b | 9.2b | 9.4ab | 9.9a | 9.3b | 0.05 | 0.01 | 0.23 | 0.77 | |
|
| 12 | 8.2b | 9.0a | 9.1a | 9.1a | 8.9a | 0.11 | <0.01 | 0.03 | 0.39 |
| 24 | 8.1 | 8.5 | 8.8 | 8.9 | 7.9 | 0.16 | 0.07 | 0.62 | 0.88 | |
| 48 | 7.7 | 8.6 | 8.5 | 8.6 | 8.4 | 0.16 | 0.10 | 0.26 | 0.46 | |
|
| 12 | 9.9 | 10.0 | 10.0 | 10.1 | 9.9 | 0.09 | 0.29 | 0.78 | 0.71 |
| 24 | 9.8 | 10.2 | 10.1 | 10.1 | 9.8 | 0.12 | 0.27 | 0.35 | 0.43 | |
| 48 | 9.7b | 10.1ab | 10.1ab | 10.4a | 9.9ab | 0.12 | 0.02 | 0.65 | 0.33 | |
|
| 12 | 10.7a | 10.2b | 9.3c | 8.3d | 8.2d | 0.09 | <0.01 | 0.03 | 0.25 |
| 24 | 9.1a | 9.2a | 8.3ab | 7.8b | 7.5b | 0.16 | 0.01 | 0.32 | 0.24 | |
| 48 | 8.6a | 8.2ab | 7.7bc | 7.2c | 7.3c | 0.13 | <0.01 | 0.69 | 0.85 | |
M-LEO = Microencapsulated-lemongrass oil (% of total dry matter substrate), IT = Incubation time, SEM = Standard error of mean, L = linear, Q = Quadratic, C = Cubic. a–d Means within the same row with different letters are significantly different at p < 0.05.
DISCUSSION
Role of microencapsulation and suitability of BSF protein as wall material
Microencapsulation is an advanced technique for protecting, stabilizing, and delivering bioactive substances across various sectors, including pharmaceuticals, cosmetics, and agriculture. By entrapping active compounds within a protective matrix and controlling their release, microencapsulation improves the functional effectiveness and stability of bioactive ingredients [38]. Protein–based encapsulants are especially appealing due to their favorable functional properties, such as film-forming ability, emulsification, and water retention. Additionally, chitin–protein complexes can control EO release by creating dense structural barriers that regulate diffusion and physically restrict microbial access to the encapsulated compounds [39].
BSF protein is a sustainable, eco-friendly encapsulating material, as BSF larvae efficiently convert organic waste into high-value protein with minimal resource use and significantly lower methane emissions than traditional livestock systems [40]. In this study, M-LEO achieved an EE of 85.2%, consistent with previous findings by Phupaboon
Functional characteristics of BSF–derived encapsulants
Using BSF–derived protein as a wall material offers clear benefits for stabilizing EO. BSF larvae are high in protein and contain significant amounts of chitin, making up about 7%–26% of DM [41]. Chitin, the second-most-abundant natural polymer, and its derivative, chitosan, are well-known biopolymers with applications in the food, medical, and agricultural fields [42]. Protein–based encapsulants also exhibit excellent functionality, including water retention, emulsification, gelation, and film formation, making them highly effective for encapsulating hydrophobic BC, such as EOs [43]. Therefore, using BSF–derived proteins as encapsulating materials offers a new and promising method for stabilizing EO designed for rumen-targeted delivery.
Effects of M-LEO on gas production and nutrient degradability
Supplementation with M-LEO increased cumulative gas production, especially at a 6% inclusion level of total DM substrate. This effect may result from the combined nutritional benefits of M-LEO, where lemongrass oil provides readily available energy, and BSF protein adds additional crude protein. However, the reduction in gas production and nutrient degradability at the 8% inclusion level indicates that excessive supplementation might hinder rumen microbial activity and fermentation efficiency. Previous
At 12, 24, and 48 h of incubation, IVDMD and IVOMD were enhanced by M-LEO supplementation, with the highest improvement seen at the 6% inclusion level. The bioactive components of M-LEO, including phenolics, flavonoids, and essential oils, may stimulate rumen microbial activity and improve substrate utilization. Supporting this idea, Amin
Modulation of rumen fermentation pathways and methane mitigation
M-LEO supplementation significantly increased total VFA concentration and the molar proportion of propionate while decreasing the acetate proportion. This change in fermentation pattern may be due to the controlled, gradual release of lemongrass bioactives from the BSF protein matrix, which alters hydrogen-utilization pathways in the rumen. Increased propionate formation diverts hydrogen away from methanogenesis, thus reducing methane production [48, 49]. Similar increases in total VFA and propionate production have been reported by Hassan
In the present study, methane production was significantly decreased at 12, 24, and 48 h following M-LEO supplementation. Suppressing ruminal methanogenesis promotes a metabolic shift from acetate-to-propionate synthesis, as propionate formation is thermodynamically favored over methane production when hydrogen availability is limited [53]. Consistent with these findings, Amin
Rumen pH, nitrogen metabolism, and microbial stability
Rumen pH and ammonia-nitrogen (NH2-N) concentrations remained unaffected by M-LEO supplementation and stayed within optimal physiological ranges for microbial activity. The observed pH range (6.87–6.94) is ideal for fibrolytic bacteria and supports efficient fiber breakdown without increasing the risk of ruminal acidosis [54]. Stable pH conditions may also help promote favorable fermentation outcomes by preserving an optimal environment for microbial metabolism [55]. Similarly, NH2-N concentrations ranged from 16.0 to 18.6 mg/dL, which aligns with the optimal levels (15–30 mg/dL) needed to support maximum microbial protein synthesis and rumen fermentation efficiency [56].
Influence of M-LEO on rumen microbial populations
Supplementation with M-LEO at 6% of total DM substrate significantly increased the abundance of key cellulolytic bacteria, including
Additionally, M-LEO increased populations of
CONCLUSION
This study showed that M-LEO, using BSF protein as a new wall material, effectively improved
The findings emphasize the potential of BSF protein–based microencapsulation as a sustainable and effective delivery system for EO in ruminant nutrition. By allowing controlled release of BC, M-LEO provides a promising feed additive strategy to enhance feed utilization efficiency and reduce enteric methane emissions, supporting climate-smart, environmentally responsible livestock production. Using insect-derived proteins also aligns with circular bioeconomy principles by transforming organic waste into high-value functional ingredients.
A key strength of this study is its innovative combination of insect biotechnology and phytogenic feed additives. Using BSF protein as an encapsulating material offers a new, sustainable alternative to traditional wall materials. Additionally, the thorough evaluation of fermentation kinetics, nutrient degradability, microbial populations, and methane production provides solid mechanistic insight into how M-LEO works.
The main limitation of this study is its
Future research should focus on validating these findings
In conclusion, BSF protein–based M-LEO is a new, sustainable, and effective method for influencing rumen fermentation and reducing methane emissions. This approach holds significant potential for developing next-generation feed additives that promote productive, environmentally friendly ruminant production systems.
DATA AVAILABILITY
All the generated data are included in the manuscript.
AUTHORS’ CONTRIBUTIONS
MM, SP, SK, TH, and MW: Conceptualization and design of the study. MM, CS, GD, RP, SP, and SS: Sampling and chemical analysis. MM: Statistical analysis. MM, SK, and MW: Drafted and revised the manuscript. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors thank the Tropical Feed Resources Research and Development Center, Department of Animal Science, Faculty of Agriculture, Khon Kaen University, Thailand, for their invaluable support and contributions. The National Science Research and Innovation Fund (NSRF) provided funding support for the research conducted on this study (Fundamental Fund; approval number 68A103000138).
REFERENCES
- Jafari S, Ebrahimi M, Goh YM, Rajion MA, Jahromi MF, Al-Jumaili WS. Manipulation of rumen fermentation and methane gas production by plant secondary metabolites (saponin, tannin and essential oil)–a review of ten-year studies. Ann Anim Sci 2019;19(1):3-29. [Google Scholar] | [Crossref]
- Kholif AE, Olafadehan OA. Essential oils and phytogenic feed additives in ruminant diet:chemistry, ruminal microbiota and fermentation, feed utilization and productive performance. Phytochem Rev 2021;20(6):1087-1108. [Google Scholar] | [Crossref]
- Benchaar C, Greathead H. Essential oils and opportunities to mitigate enteric methane emissions from ruminants. Anim Feed Sci Technol 2011;166:338-355. [Google Scholar] | [Crossref]
- Matte EHC, Luciano FB, Evangelista AG. Essential oils and essential oil compounds in animal production as antimicrobials and anthelmintics:an updated review. Anim Health Res Rev 2023;24(1):1-11. [Google Scholar] | [Crossref]
- Noshirvani N. Essential oils as natural food preservatives:special emphasis on antimicrobial and antioxidant activities. J Food Qual 2024;2024(1):5807281. [Google Scholar] | [Crossref]
- Phupaboon S, Muslykhah U, Suriyapha C, Sommai S, Matra M, Dagaew G, Prachumchai R, Wanapat M. Encapsulated phytogenic oils enhance
in vitro rumen fermentation and reduce methane emissions. BMC Vet Res 2025;21((1)):352. [Google Scholar] | [Crossref] - Abdulazeez MA, Abdullahi AS, James BD, Preedy VR. Lemongrass (
Cymbopogon spp.) oils. San Diego (CA): Academic Press; 2016. p. 509-516. [Google Scholar] - Mukarram M, Choudhary S, Khan MA, Poltronieri P, Khan MMA, Ali J, Kurjak D, Shahid M. Lemongrass essential oil components with antimicrobial and anticancer activities. Antioxidants 2022;11(1):20. [Google Scholar] | [Crossref]
- Kholif AE, Matloup OH, Hadhoud FI, Kassab AY, Adegbeye MJ, Hamdon HA. Lemongrass supplementation to Farafra ewes improved feed utilization, lactational performance and milk nutritive value in the subtropics. Anim Biotechnol 2022;33(6):1118-1127. [Google Scholar] | [Crossref]
- Peng X, Ren W, Jia M, Zhou Q, Li B, Li G, Xie Y, Dai X, Cao H, Shi X. Multi-scale study on the volatility of essential oil and its modulation methods. J Drug Deliv Sci Technol 2025;107:107227. [Google Scholar] | [Crossref]
- Al-Maqtari QA, Rehman A, Mahdi AA, Al-Ansi W, Wei M, Yanyu Z, Phyo HM, Galeboe O, Yao W. Application of essential oils as preservatives in food systems:challenges and future prospectives–a review. Phytochem Rev 2022;21(4):1209-1246. [Google Scholar] | [Crossref]
- Wang H, Wen L, Jiang F, Ren P, Yang Y, Song S, Yang Z, Wang Y. A comprehensive review of advances in hepatocyte microencapsulation:selecting materials and preserving cell viability. Front Immunol 2024;15:1385022. [Google Scholar] | [Crossref]
- Calvo P, Castaño AL, Hernández MT, González-Gómez D. Effects of microcapsule constitution on the quality of microencapsulated walnut oil. Eur J Lipid Sci Technol 2011;113:1273-1280. [Google Scholar] | [Crossref]
- Nezamdoost-Sani N, Amiri S, Khaneghah AM. The application of the coacervation technique for microencapsulation bioactive ingredients:a critical review. J Agric Food Res 2024;18:101431. [Google Scholar] | [Crossref]
- Yang X, Liang Y, Li K, Hu Q, He J, Xie J. Advances in microencapsulation of flavor substances:preparation techniques, wall material selection, characterization methods, and applications. J Agric Food Chem 2025;73(16):9459-9477. [Google Scholar] | [Crossref]
- Muslykhah U, Phupaboon S, Suriyapha C, Matra M, Wanapat M. Encapsulation of protein-based bioactive from black soldier fly for ruminant feeding. J Agric Food Res 2024:101325. [Google Scholar] | [Crossref]
- Wang X, Ni X, Duan C, Li R, Jiang XE, Xu M, Yu R. The effect of ultrasound treatment on the structural and functional properties of
Tenebrio molitor myofibrillar protein. Foods 2024;13(17):2817. [Google Scholar] | [Crossref] - Miron L, Montevecchi G, Bruggeman G, Macavei LI, Maistrello L, Antonelli A, Thomas M. Functional properties and essential amino acid composition of proteins extracted from black soldier fly larvae reared on canteen leftovers. Innov Food Sci Emerg Technol 2023;87:103407. [Google Scholar] | [Crossref]
- Bakry AM, Abbas S, Ali B, Majeed H, Abouelwafa MY, Mousa A, Liang L. Microencapsulation of oils:a comprehensive review of benefits, techniques, and applications. Compr Rev Food Sci Food Saf 2016;15(1):143-182. [Google Scholar] | [Crossref]
- Official methods of analysis. Gaithersburg (MD): Association of Official Analytical Chemists; 2012. [Google Scholar]
- Van Soest PV, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J Dairy Sci 1991;74(10):3583-3597. [Google Scholar] | [Crossref]
- Laosirisathian N, Saenjum C, Sirithunyalug J, Eitssayeam S, Sirithunyalug B, Chaiyana W. The chemical composition, antioxidant and anti-tyrosinase activities, and irritation properties of Sripanya
Punica granatum peel extract. Cosmetics 2020;7(1):1-13. [Google Scholar] | [Crossref] - Sembiring EN, Elya B, Sauriasari R. Phytochemical screening, total flavonoid and total phenolic content and antioxidant activity of different parts of
Caesalpinia bonduc (L.). Roxb. Phcog J 2018;10(1):123-127. [Google Scholar] | [Crossref] - Brand-Williams W, Cuvelier ME, Berset C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci Technol 1995;28:25-30. [Google Scholar] | [Crossref]
- Dudonné S, Vitrac X, Coutière P, Woillez M, Mérillon JM. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD and ORAC assays. J Agric Food Chem 2009;57:1768-1774. [Google Scholar] | [Crossref]
- Benzie IFF, Strain JJ. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power:the FRAP assay. Anal Biochem 1996;239:70-76. [Google Scholar] | [Crossref]
- Phupaboon S, Matra M, Prommachart R, Totakul P, Supapong C, Wanapat M. Extraction, characterization, and chitosan microencapsulation of bioactive compounds from
Cannabis sativa L., Cannabis indica L., andMitragyna speciosa K. Antioxidants 2022;11(11):2103. [Google Scholar] | [Crossref] - Menke KH, Steingass H. Estimation of the energetic feed value obtained from chemical analysis and
in vitro gas production using rumen fluid. Anim Res Dev 1988;28:7-55. [Google Scholar] | [Crossref] - Ørskov ER, McDonald I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J Agric Sci 1979;92:499-503. [Google Scholar] | [Crossref]
- Pattra S, Sangyoka S, Boonmee M, Reungsang A. Bio-hydrogen production from the fermentation of sugarcane bagasse hydrolysate by
Clostridium butyricum . Int J Hydrog Energy 2008;33(19):5256-5265. [Google Scholar] | [Crossref] - Tilley JMA, Terry DR. A two-stage technique for the
in vitro digestion of forage crops. Grass Forage Sci 1963;18(2):104-111. [Google Scholar] | [Crossref] - Viennasay B, Wanapat M, Totakul P, Phesatcha B, Ampapon T, Cherdthong A. Effect of
Flemingia macrophylla silage onin vitro fermentation characteristics and reduced methane production. Anim Prod Sci 2020;60:1918. [Google Scholar] | [Crossref] - Patra AK, Saxena J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J Sci Food Agric 2011;91:24-37. [Google Scholar] | [Crossref]
- Carrasco JMD, Cabral C, Redondo LM, Viso NDP, Colombatto D, Farber MD, Fernandez Miyakawa ME. Impact of chestnut and quebracho tannins on rumen microbiota of bovines. Biomed Res Int 2017;2017:9610810. [Google Scholar] | [Crossref]
- Bhatta R, Uyeno Y, Tajima K, Takenaka A, Yabumoto Y, Nonaka I, Enishi O, Kurihara M. Difference in the nature of tannins on
in vitro ruminal methane and volatile fatty acid production and on methanogenic archaea and protozoal populations. J Dairy Sci 2009;92:5512-5522. [Google Scholar] | [Crossref] - Becker PM, van Wikselaar PG, Franssen MCR, de Vos RCH, Hall RD, Beekwilder J. Evidence for a hydrogen-sink mechanism of (+)-catechin-mediated emission reduction of the ruminant greenhouse gas methane. Metabolomics 2014;10:179-189. [Google Scholar] | [Crossref]
- Whelan JA, Russell NB, Whelan MA. A method for the absolute quantification of cDNA using real-time PCR. J Immunol Methods 2003;278:261-269. [Google Scholar] | [Crossref]
- Lobel BT, Baiocco D, Al-Sharabi M, Routh AF, Zhang Z, Cayre OJ. Current challenges in microcapsule designs and microencapsulation processes:a review. ACS Appl Mater Interfaces 2024;16((31)):40326-40355. [Google Scholar] | [Crossref]
- Jiang L, Wang F, Xie X, Xie C, Li A, Xia N, Gong X, Zhang H. Development and characterization of chitosan/guar gum active packaging containing walnut green husk extract and its application on fresh-cut apple preservation. Int J Biol Macromol 2022;209:1307-1318. [Google Scholar] | [Crossref]
- Bukchin-Peles S, Lozneva KB, Tomberlin JK, Zilberman D. From waste management to protein innovation:black soldier fly as an embodiment of the circular bioeconomy. Future Foods 2025;11:100592. [Google Scholar] | [Crossref]
- Zhou M, Peng YJ, Chen Y, Klinger CM, Oba M, Liu JX. Assessment of microbiome changes after rumen transfaunation:implications on improving feed efficiency in beef cattle. Microbiome 2018;6:1-14. [Google Scholar] | [Crossref]
- Kumirska J, Czerwicka M, Kaczyński Z, Bychowska A, Brzozowski K, Thöming J, Stepnowski P. Application of spectroscopic methods for structural analysis of chitin and chitosan. Mar Drugs 2010;8:1567-1636. [Google Scholar] | [Crossref]
- Nesterenko A, Alric I, Silvestre F, Durrieu V. Vegetable proteins in microencapsulation:a review of recent interventions and their effectiveness. Ind Crops Prod 2013;42:469-479. [Google Scholar] | [Crossref]
- Patra AK, Yu Z. Effects of adaptation of
in vitro rumen culture to garlic oil, nitrate, and saponin and their combinations on methanogenesis, fermentation, and abundances and diversity of microbial populations. Front Microbiol 2015;6:1434. [Google Scholar] | [Crossref] - Cobellis G, Trabalza-Marinucci M, Marcotullio MC, Yu Z. Evaluation of different essential oils in modulating methane and ammonia production, rumen fermentation, and rumen bacteria
in vitro . Anim Feed Sci Technol 2016;215:25-36. [Google Scholar] | [Crossref] - Amin N, Tagliapietra F, Arango S, Guzzo N, Bailoni L. Free and microencapsulated essential oils incubated
in vitro :ruminal stability and fermentation parameters. Animals 2021;11(1):180. [Google Scholar] | [Crossref] - Muslykhah U, Phupaboon S, Suriyapha C, Sommai S, Pongsub S, Dagaew G, Matra M, Wanapat M. Effects of phytonutrient-based encapsulation of
Wolffia globosa on gas production,in vitro fermentation characteristics, and methane mitigation usingin vitro study techniques. Ital J Anim Sci 2024;23(1):1869-1883. [Google Scholar] | [Crossref] - Patra AK, Saxena J. The effect and mode of action of saponins on the microbial populations and fermentation in the rumen and ruminant production. Nutr Res Rev 2009;22:204-219. [Google Scholar] | [Crossref]
- Jalal H, Sucu E, Cavallini D, Giammarco M, Akram MZ, Karkar B, Gao M, Pompei L, Eduardo J, Prasinou P, Fusaro I. Rumen fermentation profile and methane mitigation potential of mango and avocado byproducts as feed ingredients and supplements. Sci Rep 2025;15(1):16164. [Google Scholar] | [Crossref]
- Hassan FU, Arshad MA, Ebeid HM, Rehman MSU, Khan MS, Shahid S, Yang C. Phytogenic additives can modulate rumen microbiome to mediate fermentation kinetics and methanogenesis through exploiting diet–microbe interaction. Front Vet Sci 2020;7:575801. [Google Scholar] | [Crossref]
- Suriyapha C, Phupaboon S, Dagaew G, Sommai S, Matra M, Prachumchai R, Haitook T, Wanapat M.
In vitro fermentation end-products and rumen microbiome as influenced by microencapsulated phytonutrient pellets (LEDRAGON) supplementation. Sci Rep 2024;14(1):14425. [Google Scholar] | [Crossref] - Phupaboon S, Matra M, Sommai S, Dagaew G, Suriyapha C, Prachumchai R, Wanapat M. Microencapsulation efficiency of fruit peel phytonutrient-based antimicrobial to mitigate rumen emission using
in vitro fermentation technique. Ital J Anim Sci 2024;23(1):664-677. [Google Scholar] | [Crossref] - Wang K, Xiong B, Zhao X. Could propionate formation be used to reduce enteric methane emission in ruminants?. Sci Total Environ 2023;855:158867. [Google Scholar] | [Crossref]
- Wanapat M. Manipulation of cassava cultivation and utilization to improve protein to energy biomass for livestock feeding in the tropics. Asian-Australas J Anim Sci 2003;16(3):463-472. [Google Scholar] | [Crossref]
- Kang S, Wanapat M, Phesatcha K, Norrapoke T, Foiklang S, Ampapon T, Phesatcha B. Using krabok (
Irvingia malayana ) seed oil andFlemingia macrophylla leaf meal as a rumen enhancer in anin vitro gas production system. Anim Prod Sci 2017;57(2):327-333. [Google Scholar] | [Crossref] - Wanapat M, Pimpa O. Effect of ruminal NH3-N levels on ruminal fermentation, purine derivatives, digestibility and rice straw intake in swamp buffaloes. Asian-Australas J Anim Sci 1999;12(6):904-907. [Google Scholar] | [Crossref]
- Huang H, Szumacher-Strabel M, Patra AK, Ślusarczyk S, Lechniak D, Vazirigohar M, Varadyova Z, Kozłowska M, Cieślak A. Chemical and phytochemical composition,
in vitro ruminal fermentation, methane production, and nutrient degradability of fresh and ensiledPaulownia hybrid leaves. Anim Feed Sci Technol 2021;279:115038. [Google Scholar] | [Crossref] - Agarwal N, Shekhar C, Kumar R, Chaudhary LC, Kamra DN. Effect of peppermint (
Mentha piperita ) oil onin vitro methanogenesis and fermentation of feed with buffalo rumen liquor. Anim Feed Sci Technol 2009;148:321-327. [Google Scholar] | [Crossref] - Wallace RJ, Chaudhary LC, McKain N, McEwan NR, Richardson AJ, Vercoe PE, Walker ND, Paillard D.
Clostridium proteoclasticum :a ruminal bacterium that forms stearic acid from linoleic acid. FEMS Microbiol Lett 2006;265:195-201. [Google Scholar] | [Crossref] - Maia MRG, Chaudhary LC, Bestwick SS, Richardson AJ, McKain N, Larson TR, Graham IA, Wallace RJ. Toxicity of unsaturated fatty acids to the biohydrogenating ruminal bacterium
Butyrivibrio fibrisolvens . BMC Microbiol 2010;10:52. [Google Scholar] | [Crossref] - Banik BK, Durmic Z, Erskine W, Revell CK, Vadhanabhuti J, McSweeney CS, Padmanabha J, Flematti GR, Algreiby AA, Vercoe PE. Bioactive fractions from the pasture legume
Biserrula pelecinus L. have an anti-methanogenic effect against key rumen methanogens. Anaerobe 2016;39:173-182. [Google Scholar] | [Crossref] - Hernandez A, Kholif AE, Lugo-Coyote R, Elghandour MMY, Cipriano M, Rodríguez GB, Odongo NE, Salem AZM. The effect of garlic oil, xylanase enzyme and yeast on biomethane and carbon dioxide production from 60-d-old Holstein dairy calves fed a high-concentrate diet. J Clean Prod 2017;142:2384-2392. [Google Scholar] | [Crossref]
- Prachumchai R, Suriyapha C, Dagaew G, Sommai S, Matra M, Phupaboon S, Phasuk Y, Wanapat M. Microencapsulation of lemongrass and mangosteen peel as phytogenic compounds to gas kinetics, fermentation, degradability, methane production, and microbial population using
in vitro gas technique. PLoS One 2024;19(6):e0304282. [Google Scholar] | [Crossref]