


ABSTRACT
Background and Aim: Sex sorting of sperm is a valuable reproductive biotechnology for improving herd productivity and genetic management in livestock. Recent advances have explored Toll-like receptor 7/8 (TLR7/8) activation using resiquimod (R848) as a biological approach to selectively modulate sperm function. However, its application in swamp buffalo remains limited. This study aimed to evaluate the effectiveness of R848 combined with discontinuous Percoll density gradient centrifugation (PDGC) for enriching Y-bearing sperm in frozen–thawed swamp buffalo semen and to determine the optimal concentration to maximize enrichment without compromising sperm quality.
Materials and Methods: Frozen–thawed semen from five swamp buffalo bulls (
Results: Quantitative polymerase chain reaction analysis demonstrated that treatment with 0.03 μM R848 resulted in a significant increase in BRY4a expression (approximately 2.6-fold) compared to other groups (p < 0.05), indicating enhanced Y-chromosome enrichment. Sperm motility parameters showed no significant differences in total and progressive motility between treated groups (0, 0.03, and 0.3 μM) and post-thawed controls (p > 0.05). However, velocity-related parameters such as average path velocity and curvilinear velocity increased significantly, while linearity decreased, reflecting a hyperactivation-like motility pattern. In contrast, the highest concentration (1 μM) negatively affected overall sperm motility and kinematic performance.
Conclusion: The combination of low-dose R848 (0.03 μM) and PDGC effectively enhances the relative proportion of Y-bearing sperm in frozen–thawed swamp buffalo semen without detrimental effects on motility. This approach represents a cost-effective and field-applicable alternative to conventional sperm sexing methods, with potential applications in buffalo breeding and conservation programs. Further validation using larger sample sizes and fertility trials is recommended.
Keywords: buffalo breeding, density gradient centrifugation, frozen–thawed semen, R848, sperm motility, sperm sexing, swamp buffalo, Y-chromosome enrichment.
INTRODUCTION
During the past decades,
Spermatogenesis generates two distinct populations of spermatozoa, namely X- and Y-chromosome-bearing sperm, such that male gametes determine the sex of the offspring during fertilization. Several sperm sexing techniques have been developed, including density gradient centrifugation [3], immunological-based selection [4–6], magnetically activated cell sorting (MACS) [6], swim-up [7, 8], albumin column separation [9, 10], and flow cytometry [11, 12]. Among these technologies, flow cytometry remains the gold standard, achieving up to 90% purity in separating X- and Y-chromosome-bearing sperm based on DNA content [13, 14]. However, its implementation is constrained by the requirement for skilled personnel and sophisticated instrumentation [15, 16]. In addition, the use of semen sorted by flow cytometry has been associated with decreased reproductive performance, including a 23% reduction in pregnancy rate and a 24% reduction in calving rate [15]. Therefore, alternative cost-effective and field-applicable methods are needed.
Recent studies have explored Toll-like receptor 7/8 (TLR7/8) activation as a novel approach to sperm sexing. The TLR7/8 agonist R848 has been successfully applied in cattle for sperm sexing. Umehara
Mechanistically, R848 selectively reduces X-sperm motility by impairing mitochondrial adenosine triphosphate (ATP) production, thereby creating motility differences that allow sperm separation [20]. This mechanism has been demonstrated in mice, cattle, goats, and sheep, where X-sperm exhibit reduced motility, facilitating preferential selection of Y-sperm [17–22]. However, the efficacy of R848-mediated sperm sexing appears to be species-dependent. A study in dogs [23] reported no significant enrichment following 0.4 μM R848 treatment for 60 min, with Y- and X-sperm proportions remaining comparable to controls. These findings indicate that species-specific variation in TLR7/8 signaling influences the effectiveness of this approach.
To enhance the separation efficiency, an additional physical selection step can be applied. Percoll-based density gradient centrifugation (PDGC) is widely used in bovine
Despite advances in sperm sexing technologies, significant limitations remain in developing a practical, efficient, and species-specific method for swamp buffalo. Current gold-standard techniques, such as flow cytometry, are costly, technically demanding, and associated with reduced fertility outcomes, limiting their field applicability. Although TLR7/8 activation with R848 has shown promising results in several livestock species, its effectiveness is highly species-dependent, and its application in swamp buffalo remains underexplored. Furthermore, previous studies have primarily focused on either biochemical modulation or physical separation techniques, with limited investigation of their combined effects. The lack of integrated approaches that simultaneously exploit molecular signaling pathways and physical sperm selection mechanisms represents a critical knowledge gap. In addition, the optimal concentration of R848 for achieving effective Y-sperm enrichment without compromising sperm motility and functional integrity in swamp buffalo has not been clearly established. Therefore, there is a need for systematic evaluation of combined strategies that are both biologically effective and practically applicable under field conditions.
The present study aimed to evaluate the effectiveness of combining resiquimod (R848), a TLR7/8 agonist, with PDGC for sex selection of sperm in swamp buffalo. Specifically, this study sought to determine the optimal concentration of R848 that maximizes Y-bearing sperm enrichment while maintaining sperm motility and functional characteristics. In addition, the study aimed to assess whether the integration of molecular modulation through TLR7/8 activation and physical separation via PDGC could provide a cost-effective and field-applicable alternative to conventional sperm sexing techniques. The findings are expected to contribute to the development of practical strategies to improve reproductive efficiency, support genetic selection, and enhance the sustainability of swamp buffalo breeding and conservation programs.
MATERIALS AND METHODS
Ethical approval
This study was conducted in accordance with the ethical guidelines and regulatory standards for the use of animals in research. The experimental protocol was reviewed and approved by the Institutional Animal Care and Use Committee of Khon Kaen University, Thailand, under Approval Record No. IACUC-KKU (C)-112/67, Reference No. 660201.2.11/820 (60), dated October 25, 2024.
Frozen semen samples from mature swamp buffalo bulls were obtained from the Department of Livestock Development, Thailand. No live animals were handled, restrained, subjected to experimental procedures, or exposed to any intervention specifically for this study. Therefore, the study involved only the laboratory use of cryopreserved semen samples supplied by an authorized governmental source. All procedures involving semen handling, thawing, incubation, PDGC, sperm motility assessment, cryopreservation, DNA extraction, and quantitative polymerase chain reaction analysis were performed under approved laboratory conditions, in accordance with institutional biosafety and animal research guidelines.
Study period and location
This study was conducted during 2024 at the Faculty of Veterinary Medicine, Khon Kaen University, Khon Kaen, Thailand. Laboratory analyses, including sperm processing and molecular evaluation, were performed in associated research laboratories equipped for reproductive biotechnology and molecular biology.
Study design
Frozen semen from five mature swamp buffalo bulls, aged 4–12 years, was obtained from the Department of Livestock Development (Thailand). Frozen–thawed sperm were incubated with R848 at concentrations of 0, 0.03, 0.3, and 1 μM. Following incubation, sperm were subjected to PDGC to separate subpopulations based on physical and functional characteristics. Sperm collected from the pellet fraction were evaluated for motility using computer-assisted sperm analysis (CASA), while the remaining sperm were re-frozen for sex determination using real-time polymerase chain reaction (PCR).
Sperm sex was determined using real-time PCR with two primer sets: BRY4a, a Y-chromosome-specific marker, and GAPDH, used as an internal housekeeping gene control. Each sample was analyzed in triplicate.
Reagent and media preparation
All reagents and chemicals were purchased from Sigma Chemical Company (St. Louis, MO, USA). The PDGC solution (Percoll™ Plus, Lot No. 10326907) was obtained from Cytiva (Uppsala, Sweden). Real-time PCR reagents were sourced from Qiagen (USA). Primers were supplied by Integrated DNA Technologies (IDT, USA).
Semen preparation and sperm incubation
Sixteen frozen sperm straws cryopreserved in a 20% egg yolk–Tris (EYT) extender consisting of 250 mmol/L Tris (Tris-hydroxymethyl aminomethane), 90 mmol/L citric acid, 70 mmol/L fructose, 100 IU/mL penicillin G, 100 μg/mL streptomycin, 20% (v/v) fresh hen egg yolk, and 7% (v/v) glycerol were thawed at 37°C for 30 s. Immediately after thawing, sperm motility characteristics were assessed using CASA.
Thawed semen was aliquoted to 1 mL (10 × 107 spermatozoa) and resuspended in 2 mL of Tris-based buffer (Tris 15.4 g, citric acid 8.65 g, glucose 6.25 g) supplemented with R848 (TLR7/8 ligand; SML0196-10MG, Sigma-Aldrich, St. Louis, MO, USA) to obtain final concentrations of 0 μM (control), 0.03 μM, 0.3 μM, and 1 μM. The 0 μM group served as a process-matched control, whereas the post-thawed group represented the untreated baseline. Samples were incubated at 37°C for 45 min in a humidified atmosphere containing 5% CO2.
Sperm selection through discontinuous PDGC
Sperm separation was performed using a modified Percoll density gradient method adapted [32]. A 90% (v/v) Percoll solution was prepared by combining Percoll™ Plus (Cytiva, Uppsala, Sweden) with 10× Tyrode’s Albumin Lactate Pyruvate–HEPES (TALP-HEPES) buffer (9:1 ratio). The TALP-HEPES buffer consisted of sodium chloride, potassium chloride, sodium dihydrogen phosphate, and HEPES, supplemented with calcium chloride, magnesium chloride, and lactic acid, with pH adjusted to 7.3. A 45% Percoll solution was prepared by diluting the 90% Percoll solution with TALP-HEPES (1:1 ratio).
After incubation, 3 mL of semen from each group was layered onto a discontinuous Percoll gradient (2 mL 45% over 2 mL 90%) in a 15 mL conical tube and centrifuged at 590 g for 10 min (Hermle Z 320 centrifuge, Hermle Labortechnik, Wehingen, Germany). The bottom pellet (~1 mL), expected to be enriched in Y-bearing spermatozoa, was recovered and washed in 5 mL TALP-HEPES by centrifugation at 330 g for 7 min. The pellet was resuspended in TALP to a final volume of 500 μL and loaded into 0.5 mL straws for refreezing using EYT extender.
Sperm freezing and thawing procedure
Sperm suspensions were loaded into 0.5 mL plastic straws and equilibrated in liquid nitrogen vapor at approximately 10 cm above the liquid nitrogen surface for 10 min. Following vapor-phase equilibration, the straws were plunged directly into liquid nitrogen and stored in cryogenic containers until further use.
Assessment of sperm motility using CASA
Sperm motility characteristics were assessed using CASA (CEROS II system, Hamilton Thorne, Beverly, MA, USA; software version 1.14 Rev.A). After thawing at 37°C for 30 s, 15 μL of semen was placed in Leja chamber slides (20 μm; SC 20-01-C, Leja Products B. V., Nieuw-Vennep, The Netherlands) and incubated at 37°C for 5 min before analysis.
Samples were analyzed using a Nikon 10× objective (Nikon, Japan) with calibration factors of 1.21 for both X and Y axes. Videos were recorded at 37°C with a frame capture speed of 60 Hz and 30 frames per field. A minimum of 200 spermatozoa were analyzed per sample.
Cell detection parameters were defined as follows: head size 10–50 μm², head brightness ≥170, elongation 1–100%, tail brightness ≥70, and minimum large object area 1000 μm². Progressive motility was defined as average path velocity (VAP) ≥50 μm/s and STR ≥80%. Slow sperm were defined as VAP ≥20 μm/s and VSL ≥30 μm/s, whereas static sperm were defined as VAP ≤4 μm/s and VSL ≤1 μm/s.
Motility parameters included curvilinear distance (DCL), straight-line distance (DSL), average path distance (DAP), curvilinear velocity (VCL), VAP, straight-line velocity (VSL), straightness (STR), linearity (LIN), wobble (WOB), beat cross frequency (BCF), and amplitude of lateral head displacement (ALH).
Quantification of Y-chromosome marker by real-time PCR
DNA sample preparation and extraction: Four 0.5 mL straws were thawed at 37°C for 30 s. Samples were transferred to 1.5 mL microcentrifuge tubes and centrifuged at 6010 g for 1 min. The supernatant was discarded, retaining approximately 100 μL of sperm pellet.
Sperm lysis was performed using a buffer containing Tris-HCl (pH 8.0), EDTA, sodium chloride, SDS, Proteinase K, and dithiothreitol at 55°C for 2 h. Genomic DNA was extracted using the DNeasy® Blood & Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. DNA was eluted in 80 μL AE buffer. DNA concentration and purity were measured using a BioDrop Duo spectrophotometer (BioDrop Ltd., Cambridge, UK), and samples were stored at −80°C until analysis.
Quantitative real-time PCR assay: Quantitative PCR (qPCR) was performed to determine the relative abundance of Y-chromosome-specific DNA sequences. Each 20 μL reaction contained 2 μL template DNA, ThunderBird® Next SYBR® qPCR Mix (Toyobo Co., Ltd., Osaka, Japan), and 0.4 μM of each primer. The gene BRY4a was used as the target, and GAPDH as the endogenous reference gene (Table 1).
Table 1. Primers used for real-time polymerase chain reaction.
| Primer | Sequence | Annealing temperature (°C) | Fragment length | Reference |
|---|---|---|---|---|
| BRY4a | F: 5’-CTCAGCAAAGCACACCAGAC-3’ | 56 | 300 bp | [7] |
| R: 5’-GAACTTTCAAGCAGCTGAGGC-3’ | ||||
| GAPDH | F: 5’-ACA CCC TCA AGA TTG TCA GCA A-3’ | 55–56 | 102 bp | [25] |
| R: 5’-TCA TAA GTC CCT CCA CGA TGC-3’ |
Reactions were conducted in triplicate using a CFX96 real-time PCR detection system (Bio-Rad Laboratories, Hercules, CA, USA). Negative controls without template DNA were included. The thermal cycling protocol consisted of an initial denaturation at 95°C for 2 min, followed by 40 cycles of 95°C for 5 s, 58°C for 30 s, and 72°C for 30 s, with a final extension at 65°C for 5 s.
Relative gene expression was calculated using the 2^−ΔΔCT method, normalized to GAPDH expression [7]. Melting curve analysis confirmed amplification specificity, with a single peak observed for both BRY4a and GAPDH.
Statistical analysis
All data are expressed as mean ± standard error of the mean (SEM). Data were assessed for normality and homogeneity of variance before analysis. Mean relative fold change, motility, and kinematic parameters were analyzed using one-way analysis of variance, with bulls (n = 5) treated as blocks and R848 concentrations as fixed factors.
Post hoc comparisons were performed using Duncan’s multiple range test when significance was detected (p < 0.05). Statistical analyses were conducted using SPSS software (SPSS Inc., Chicago, IL, USA), and graphs were generated using GraphPad Prism Version 9.3.0 (www.graphpad.com).
RESULTS
Assessment of sperm sex by real-time PCR
Quantitative real-time PCR analysis was conducted to assess the relative fold changes in BRY4a expression in sperm recovered from the pellet fraction following R848 treatment. Untreated spermatozoa served as the calibrator sample. Data were normalized to the endogenous reference gene GAPDH, and relative quantification was calculated using the 2^−ΔΔCt method. In the untreated control, ΔCt was defined as zero, yielding a relative expression value of 1.00. For treated samples, the 2^−ΔΔCt value reflected the fold change in BRY4a expression relative to the control [7].
The mean relative expression of BRY4a in semen treated with 0.03 μM R848 followed by PDGC showed a significantly higher value (p < 0.05) than the other groups (Table 2)., with an average 2^−ΔΔCt value corresponding to a 2.6-fold increase (Figure 1).
Table 2. Relative BRY4a expression (2^−ΔΔCt, mean ± SEM) in the pellet fraction following treatment with different concentrations of R848.
| Concentration of R848 | BRY4a (mean ± SEM) |
|---|---|
| Post-thawed | 1.00 ± 0.23ᵃ |
| 0 μM | 1.00 ± 0.03ᵃ |
| 0.03 μM | 2.64 ± 0.83ᵇ |
| 0.3 μM | 0.88 ± 0.07ᵃ |
| 1 μM | 1.17 ± 0.20ᵃ |
Values are expressed relative to the control and represent mean ± SEM. Different superscript letters within the BRY4a column indicate statistically significant differences among treatments (p < 0.05).

Figure 1. Relative expression (2^−ΔΔCt) of the Y-chromosome marker BRY4a in the pellet fraction following incubation with different concentrations of R848 and PDGC. Different superscript letters within the BRY4a column indicate statistically significant differences among treatments (p < 0.05).
Assessment of sperm motility using CASA
Sperm motility after thawing: After thawing, the mean percentages of total motile and progressive motile spermatozoa from five bulls were 79.0 ± 2.2% and 49.6 ± 5.3%, respectively. The VAP, VSL, and VCL were 92.0 ± 4.9 μm/s, 85.5 ± 4.9 μm/s, and 137.8 ± 5.4 μm/s, respectively. In addition, STR, LIN, and amplitude of lateral head displacement (ALH) were 93.0 ± 0.5%, 63.3 ± 1.7%, and 4.9 ± 0.4 μm, respectively.
Sperm motility after R848 treatment followed by PDGC: After treatment, sperm from the pellet fraction were resuspended and evaluated using CASA. The sperm motility patterns are presented in Table 3. Sperm recovery rates from the pellet fraction were 59.58 ± 15.67% (0 μM), 63.91 ± 13.17% (0.03 μM), 62.62 ± 9.64% (0.3 μM), and 56.76 ± 11.87% (1 μM).
Table 3. Motility characteristics of post-thawed buffalo sperm and post-treatment with different concentrations of R848 followed by Percoll density gradient centrifugation.
| Treatment | R848 concentration | Motility (%) | Progressive (%) | VAP (μm/s) | VSL (μm/s) | VCL (μm/s) | STR (%) | LIN (%) | ALH (μm) | WOB (%) | BCF (Hz) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Post-thawed | – | 78.98 ± 2.24ᵃ | 49.64 ± 5.25ᵃ | 92.03 ± 4.91ᵃ | 85.52 ± 4.89ᵃ | 137.77 ± 5.41ᵃ | 92.97 ± 0.52ᵃ | 63.30 ± 1.74ᶜ | 4.90 ± 0.40ᵃ | 67.85 ± 1.50ᶜᵈ | 36.36 ± 2.33ᵇ |
| Post-treatment | 0 μM | 85.88 ± 2.04ᵃ | 41.38 ± 3.09ᵃ | 115.06 ± 5.45ᵇ | 100.01 ± 5.99ᵃ | 206.80 ± 14.41ᵇ | 87.09 ± 3.06ᵃ | 50.50 ± 3.10ᵃ | 8.94 ± 0.82ᵇ | 57.58 ± 2.17ᵃ | 27.66 ± 1.72ᵃ |
| 0.03 μM | 79.86 ± 5.00ᵃ | 47.92 ± 3.50ᵃ | 115.29 ± 5.90ᵇ | 104.68 ± 5.11ᵃ | 198.42 ± 11.22ᵇ | 91.35 ± 0.54ᵃ | 55.00 ± 1.40ᵃᵇ | 8.39 ± 0.59ᵇ | 60.04 ± 1.26ᵃᵇ | 27.19 ± 0.77ᵃ | |
| 0.3 μM | 85.30 ± 1.19ᵃ | 48.10 ± 2.51ᵃ | 111.84 ± 6.67ᵇ | 102.85 ± 5.93ᵃ | 181.55 ± 13.45ᵇ | 92.44 ± 0.19ᵃ | 58.30 ± 1.53ᵇᶜ | 7.71 ± 0.58ᵇ | 63.09 ± 1.48ᵇᶜ | 26.76 ± 0.81ᵃ | |
| 1 μM | 76.80 ± 1.75ᵃ | 44.55 ± 1.68ᵃ | 97.00 ± 6.72ᵃᵇ | 90.55 ± 5.83ᵃ | 142.83 ± 12.90ᵃ | 93.57 ± 0.92ᵃ | 65.18 ± 2.83ᶜ | 5.78 ± 0.64ᵃ | 69.49 ± 2.32ᵈ | 29.16 ± 1.06ᵃ |
VAP = Average path velocity, VCL = Curvilinear velocity, VSL = Straight-line velocity, STR = Straightness, LIN = Linearity, WOB = Wobble, ALH = Amplitude of lateral head displacement, BCF = Beat cross frequency. Different superscript letters within the same column indicate significant differences among treatment groups (p < 0.05). Data are presented as mean ± standard error of the mean (SEM) from three replications.
There were no significant differences in total motility, progressive motility, VSL, and STR among treatment groups and the post-thawed group (p > 0.05). In contrast, significant increases in VAP, VCL, and ALH were observed after treatment (p < 0.05). Conversely, LIN, WOB, and BCF were significantly reduced (p < 0.05) (Table 3) (Figure 2).
DISCUSSION
Relative enrichment of Y-bearing sperm after R848 and PDGC treatment
The present study represents the first application of R848, a TLR7/8 agonist, combined with discontinuous PDGC in frozen–thawed swamp buffalo semen. The results suggest that this combined approach may increase the relative proportion of Y-bearing spermatozoa while maintaining motility parameters. Quantitative real-time PCR analysis showed that 0.03 μM R848 yielded the highest relative expression of the Y-chromosome marker BRY4a compared with the other groups. These findings indicate a relative enrichment of Y-bearing sperm at this concentration, as inferred from molecular quantification. However, the enrichment was assessed using relative qPCR analysis without absolute quantification or analytical validation of the BRY4a assay in swamp buffalo. Therefore, the findings should be interpreted as evidence of relative molecular enrichment rather than a precise determination of the proportion of Y-bearing sperm.
Sperm motility and kinematic changes after treatment
Post-thaw motility characteristics confirmed that the semen used in this study exhibited high-quality motility, indicating that sperm maintained good functional integrity before experimental treatment. Following R848 treatment, spermatozoa presumed to be less responsive to TLR7/8 activation retained progressive motility and were preferentially enriched in the pellet fraction during PDGC. This observation is consistent with previous studies showing that spermatozoa with superior motility tend to migrate into denser layers under centrifugal force [24, 25, 30–35].
Comparisons between post-thawed and post-treatment sperm (0, 0.03, and 0.3 μM) revealed distinct changes in sperm kinematic patterns following treatment, incubation, and PDGC. Velocity parameters such as VCL and VAP increased significantly, while ALH was also elevated. In contrast, LIN decreased markedly. This combined profile, characterized by high VCL and ALH with low LIN, resembles kinematic patterns associated with hyperactivation, reflecting vigorous but less linear flagellar movement [36–38]. These motility patterns are biologically relevant, as previous studies have shown that R848-induced TLR7/8 activation suppresses ATP production and motility in X-bearing sperm, whereas Y-bearing sperm remain metabolically less affected and maintain stronger propulsive capacity [17–22, 39, 40]. Therefore, the increased motility parameters observed in this study likely reflect enrichment of highly motile Y-bearing sperm within the PDGC pellet fraction. Although the observed kinematic profile is consistent with hyperactivation, definitive confirmation of hyperactivation or capacitation would require additional functional or biochemical assays.
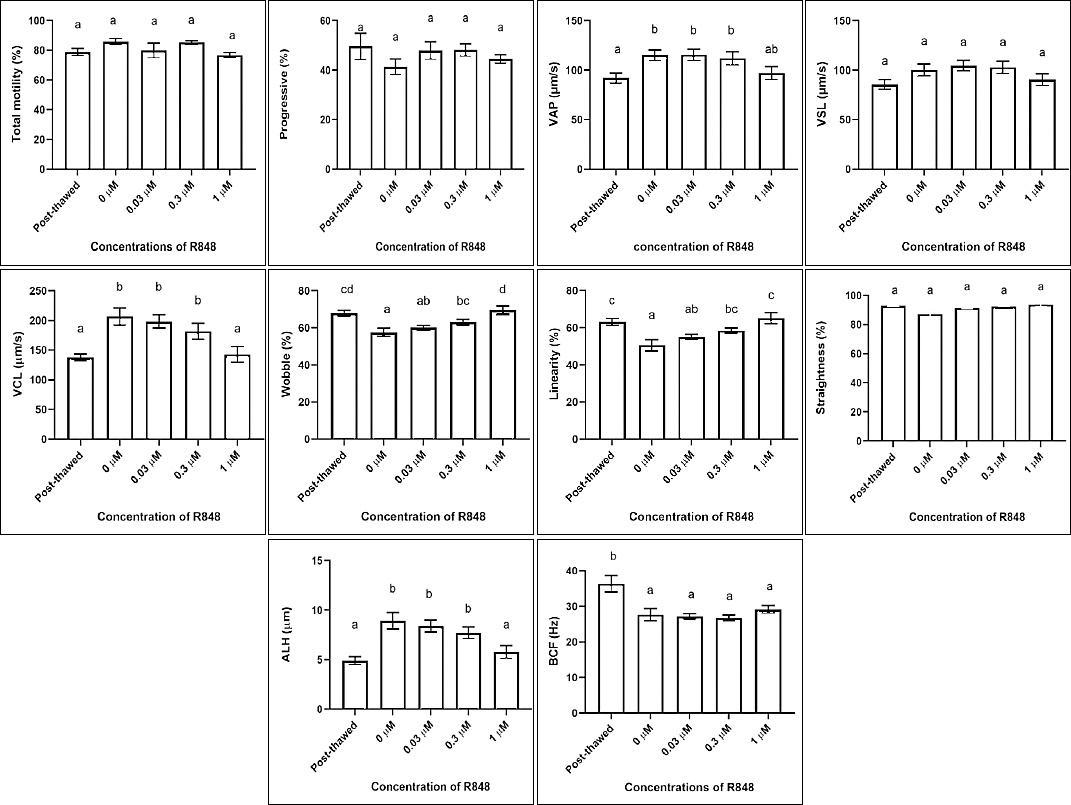
Figure 2. Motility characteristics of buffalo sperm post-thawed compared with post-treatment. Total motility = progressive motility, VAP = average path velocity, VSL = straight-line velocity, VCL = curvilinear velocity, STR = straightness, LIN = linearity (VSL/VCL × 100), ALH = amplitude of lateral head displacement, WOB = wobble (VAP/VCL × 100), BCF = beat cross frequency. Different letters above the bars indicate significant differences (p < 0.05).
Effect of high-dose R848 on sperm function
In contrast, exposure to the highest R848 concentration (1 μM) resulted in a markedly different outcome. CASA analysis showed significant reductions in total and progressive motility, as well as decreases in major velocity parameters (VCL, VAP, and VSL). Unlike the selective modulation observed at lower concentrations, 1 μM R848 appears to exert a broad inhibitory effect on overall sperm function. Excessive TLR7/8 activation at this concentration likely disrupts mitochondrial ATP synthesis and induces intracellular stress responses, impairing flagellar movement in both X- and Y-bearing spermatozoa [21]. These findings suggest that 1 μM exceeds the physiological tolerance threshold of frozen–thawed buffalo sperm, making it unsuitable for sperm sexing applications.
Comparison with previous buffalo sperm sexing studies
The findings of the present study differ from those reported by Meel
Practical implications for buffalo breeding and conservation
In summary, integration of molecular evidence with kinematic data indicates that the use of a low-dose of R848 (0.03 μM) combined with PDGC may increase the proportion of Y-bearing spermatozoa in frozen–thawed swamp buffalo semen while preserving motility parameters. This approach may provide a practical and cost-effective strategy applicable to semen stored in genetic resource banks. The technique has potential implications for conservation and breeding programs aimed at improving and managing swamp buffalo populations in Thailand. Increasing the proportion of male offspring may support agricultural labor demands and contribute to long-term sustainability of buffalo genetic resources. However, further validation in larger populations and through functional fertility assessments is necessary before routine application can be recommended.
CONCLUSION
The present study demonstrated that the application of a low concentration of R848 (0.03 μM) combined with PDGC can increase the relative abundance of Y-bearing spermatozoa in frozen–thawed swamp buffalo semen without compromising sperm motility. Molecular evaluation using qPCR targeting BRY4a revealed a significant increase in relative expression at 0.03 μM compared with other treatment groups (p < 0.05), indicating effective enrichment of Y-chromosome-bearing sperm. In parallel, CASA-based kinematic analysis showed that key motility parameters such as VAP, VCL, and ALH increased, while LIN decreased, suggesting a shift toward hyperactivation-like motility patterns that may favor the selection of highly motile Y-bearing sperm.
From a practical perspective, this combined approach provides a promising alternative to conventional sperm sexing techniques such as flow cytometry, which are often limited by high cost, technical complexity, and reduced fertility outcomes. The integration of molecular modulation through TLR7/8 activation and physical separation via PDGC offers a relatively simple, cost-effective, and field-applicable method. This strategy is particularly relevant for buffalo production systems in Thailand and similar regions, where increasing the proportion of male offspring can enhance meat production, improve breeding efficiency, and support agricultural labor demands.
A key strength of this study lies in the combined evaluation of molecular and functional parameters, linking BRY4a-based relative quantification with detailed CASA-derived kinematic analysis. This integrative approach provides a more comprehensive understanding of sperm selection dynamics compared with studies relying on a single method. In addition, the use of frozen–thawed semen enhances the practical applicability of the findings, as it reflects real-world conditions in artificial insemination and genetic resource banking systems.
However, several limitations should be acknowledged. The study relied on relative qPCR quantification without absolute validation of Y-bearing sperm proportions, thereby limiting the precise interpretation of enrichment efficiency. The sample size was relatively small, with semen obtained from only five bulls, and inter-individual variability in response to R848 treatment was not assessed. Furthermore, functional fertility outcomes, such as fertilization rates and offspring sex ratios
Future research should focus on validating these findings using larger and more diverse buffalo populations, incorporating absolute quantification techniques and advanced molecular assays. Studies assessing fertilization outcomes
In conclusion, the combined use of R848 at an optimized low concentration and PDGC represents a promising and practical strategy for enhancing enrichment of Y-bearing sperm in swamp buffalo semen. While the current findings provide strong preliminary evidence, further validation through functional and large-scale studies is required before routine application in breeding programs. This approach has the potential to contribute significantly to sustainable buffalo production, genetic improvement, and conservation efforts.
DATA AVAILABILITY
All data generated or analyzed during this study are included in this published article.
AUTHORS’ CONTRIBUTIONS
PK: Conceptualization, methodology, investigation, writing – review and editing, and supervision. NWYD: Conceptualization, methodology, investigation, and writing – original draft. PN: Methodology, investigation, and writing – review and editing. SS: Conceptualization, methodology, investigation, writing – review and editing, and supervision. ST: Sample collection and writing – review and editing. PP: Methodology and writing – review and editing. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
This research was financially supported by a research grant from the Faculty of Veterinary Medicine, Khon Kaen University (2024). Additional funding was provided by the ASEAN-GMS Scholarship (2021), Khon Kaen University. The authors gratefully acknowledge the Veterinary Diagnostic Laboratory (VDL) for providing laboratory facilities and technical assistance, and the Department of Livestock Development (DLD), Khon Kaen Province, Thailand, for supplying the semen samples used in this study.
REFERENCES
- Meepradit P, Kulwong S, Sa-ngiamsak T. Risk study on buffalo racing festival in Chonburi province. Public Health J. Burapha Univ 2008;3(2):53-58. [Google Scholar] | [Crossref]
- Malatham C, Jaruworn P. Honor, wealth and power: Folklore and identity of the people and communities in the “buffalo racing tradition”in Chonburi province. J. Thai Stud 2025;21(1):1-30. [Google Scholar] | [Crossref]
- Promthep K, Satitmanwiwat S, Kitiyanant N, Tantiwattanakul P, Jirajaroenrat K, Sitthigripong R. Practical use of Percoll density gradient centrifugation on sperm sex determination in commercial dairy farm in Thailand. Indian J. Anim. Res 2016;50(3):310-313. [Google Scholar] | [Crossref]
- Paitoon P, Sartsook A, Thongkham M, Sathanawongs A, Lumsangkul C, Pattanawong W. Sperm quality variables of sex-sorted bull semen produced by magnetic-activated cell sorting coupled with recombinant antibodies targeting Y-chromosome-bearing sperm. Theriogenology 2024;219:11-21. [Google Scholar] | [Crossref]
- Sringarm K, Thongkham M, Mekchay S, Chlumsangkul C, Thaworn W, Pattanawong W. High-efficiency bovine sperm sexing using magnetic-activated cell sorting by coupling scFv antibodies specific to Y-chromosome-bearing sperm on magnetic microbeads. Biology (Basel) 2022;11(5):715. [Google Scholar] | [Crossref]
- Phiphattanaphiphop C, Leksakul K, Wanta T, Khamlor T, Phattanakun R. Antibody-conjugated magnetic beads for sperm sexing using a multi-wall carbon nanotube microfluidic device. Micromachines 2022;13(3):1-18. [Google Scholar] | [Crossref]
- Husna AU, Awan MA, Abid Mehmood A, Sultana T, Shahzad Q, Ansari MS. Sperm sexing in Nili-Ravi buffalo through modified swim-up: Validation using SYBR®green real-time PCR. Anim. Reprod. Sci 2017;182:69-76. [Google Scholar] | [Crossref]
- Pinto S, Carrageta DF, Alves MG, Rocha A, Agarwal A, Barros A. Sperm selection strategies and their impact on assisted reproductive technology outcomes. Andrologia 2021;53(2):1-18. [Google Scholar] | [Crossref]
- Priyanto L, Herdis H, Santoso S, Anwar RI, Priyatno TP, Sitaresmi PL. The reproductive success of Simmental bovine after sex-sorting under various incubation and centrifugation protocols. Vet. World 2023;16(3):631-637. [Google Scholar] | [Crossref]
- Masrizal M, Afriani T, Wahyudi D, Saharani S, Ananda A. The quality of frozen buffalo sperm following sexing using bovine serum albumin column and swim-up methods. Bul. Peternak 2024;48(1):55-63. [Google Scholar] | [Crossref]
- Carvalho JO, Sartori R, Rodello L, Mourão GB, Bicudo SD, Margot MA. Flow cytometry sex sorting affects bull sperm longevity and compromises their capacity to bind to oviductal cells. Livest. Sci 2017;207(3):30-37. [Google Scholar] | [Crossref]
- Umezu K, Hiradate Y, Numabe T, Hara K, Tanemura K. Effects on glycocalyx structures of frozen-thawed bovine sperm induced by flow cytometry and artificial capacitation. J. Reprod. Dev 2017;63(5):473-480. [Google Scholar] | [Crossref]
- Azizeddin A, Ashkar FA, King WA, Revay T. Enrichment of Y-chromosome-bearing bull spermatozoa by swim-up through a column. Reprod. Domest. Anim 2014;49(1):e1-4. [Google Scholar] | [Crossref]
- Naniwa Y, Sakamoto Y, Toda S, Uchiyama K. Bovine sperm sex-selection technology in Japan. Reprod. Med. Biol 2019;18(1):17-26. [Google Scholar] | [Crossref]
- Reese S, Pirez MC, Steele H, Kölle S. The reproductive success of bovine sperm after sex-sorting: A meta-analysis. Sci. Rep 2021;11(1):1-18. [Google Scholar] | [Crossref]
- Steele H, Makri D, Maalouf WE, Reese S, Kölle S. Bovine sperm sexing alters sperm morphokinetics and subsequent early embryonic development. Sci. Rep 2020;10(1):1-18. [Google Scholar]
- Umehara T, Tsujita N, Zhu Z, Ikedo M, Shimada M. A simple sperm-sexing method that activates TLR7/8 on X sperm for the efficient production of sexed mouse or cattle embryos. Nat. Protoc 2020;15(8):2645-2667. [Google Scholar]
- Wen F, Liu W, Li Y, Zou Q, Xian M, Han S. TLR7/8 agonist (R848) inhibits bovine X sperm motility via PI3K/GSK3α/βand PI3K/NFκB pathways. Int. J. Biol. Macromol 2023;232:123485. [Google Scholar]
- Huang F, Niu P, Qu H, Cheng H, Wang J, Suo J. Polyvinylpyrrolidone can effectively improve the efficiency of resiquimod in sorting bovine Y sperm. Anim. Biosci 2025;38(9):1881-1893. [Google Scholar]
- Umehara T, Tsujita N, Shimada M. Activation of toll-like receptor 7/8 encoded by the X chromosome alters sperm motility and provides a novel simple technology for sexing sperm. PLoS Biol 2019;17(8):1-24. [Google Scholar]
- Ren F, Xi H, Ren Y, Li Y, Wen F, Xian M. TLR7/8 signalling affects X-sperm motility via the GSK3α/β-hexokinase pathway for the efficient production of sexed dairy goat embryos. Anim. Sci. Biotechnol 2021;12:89. [Google Scholar]
- Huang M, Cao XY, He QF, Yang HW, Chen YZ, Zhao JL. Alkaline semen diluent combined with R848 for separation and enrichment of dairy goat X-sperm. J. Dairy Sci 2022;105:10020-10032. [Google Scholar]
- Pan C, Xu S, Zhang W, Zhao Y, Zhao J, Song M. Expression of TLR7/8 in canine sperm and evaluation of the effect of ligand R848 on the sorting of canine X/Y sperm. Theriogenology 2025;231(8):127-132. [Google Scholar]
- Wolf CA, Brass KE, Rubin MIB, Pozzobon SE, Mozzaquatro FD. The effect of sperm selection by Percoll or swim-up on the sex ratio of
in vitro -produced bovine embryos. Anim. Reprod 2008;5(3/4):110-115. [Google Scholar] - Guimarães ACG, Leivas FG, Santos FW, Schwengber EB, Giotto AB, Machado CIU. Reduction of centrifugation force in discontinuous Percoll gradients increases
in vitro fertilization rates without reducing bovine sperm recovery. Anim. Reprod. Sci 2014;146(3–4):103-110. [Google Scholar] - Arias ME, Andara K, Briones E, Felmer R. Bovine sperm separation by swim-up and density gradients (Percoll and BoviPure): Effect on sperm quality, function and gene expression. Reprod. Biol 2017;17(2):126-132. [Google Scholar]
- Missio D, Folchini NP, Leivas FG, Pavin CIIUM, Pinto HF, Santos Cibin FWS. Reduction in Percoll volume increases recovery rate of sex-sorted semen of bulls without affecting sperm quality and early embryonic development. Anim. Reprod. Sci 2018;192:146-153. [Google Scholar]
- Morrell JM. Effect of colloid centrifugation on boar sperm quality during storage and function in
in vitro fertilization. Theriogenology 2019;137:122-128. [Google Scholar] - Noguchi M, Yoshioka K, Hikono H, Iwagami G, Suzuki C, Kikuchi K. Centrifugation on Percoll density gradient enhances motility, membrane integrity and
in vitro fertilizing ability of frozen-thawed boar sperm. Zygote 2013;23(2):1-8. [Google Scholar] - Fleming S, Morroll D, Nijs M. Sperm separation and selection techniques to mitigate sperm DNA damage. Life (Basel) 2025;15:302. [Google Scholar]
- Safa F, Yekti APA, Utami P, Syah HA, Febrianto N, Rachmawati A. Sperm quality, kinematics, membrane integrity, and DNA fragmentation of frozen sexed semen in Holstein-Friesian bulls. Trop. Anim. Sci. J 2025;48(3):199-210. [Google Scholar]
- Parrish JJ, Krogenaes A, Susko-Parrish JL. Effect of bovine sperm separation by either swim-up or Percoll method on success of
in vitro fertilization and early embryonic development. Theriogenology 2025;44(6):859-869. [Google Scholar] - Trigal B, Gómez E, Caamaño JN, Muñoz M, Moreno J, Carrocera S.
In vitro andin vivo quality of bovine embryos produced with sex-sorted sperm. Theriogenology 2012;78(7):1465-1475. [Google Scholar] - Akal E, Selçuk M, Esin B, Akar M, Kaya K. Comparison of the effects of different thawing methods on post-thaw sperm characteristics of buffalo bull semen. Thai J. Vet. Med 2021;51(4):697-704. [Google Scholar]
- Franchi FF, Musmeci G, Alyfanti E, Puelker R, Lodde V, Franciosi F. Optimization of
in vitro fertilization with X-sorted spermatozoa in cattle. Front. Anim. Sci 2026;6(1):1-10. [Google Scholar] - Hackerova L, Pilsova A, Pilsova Z, Zelenkova N, Hegrova PT, Klusackova B. Boar sperm motility assessment using computer-assisted sperm analysis: Current practices, limitations, and methodological challenges. Animals (Basel) 2025;15(3):1-34. [Google Scholar]
- Traini G, Ragosta ME, Tamburrino L, Papini A, Cipriani S, Vignozzi L. Microfluidic sorting can be applied for assisted reproduction sperm selection in different cases of semen abnormalities. Life (Basel) 2025;15:790. [Google Scholar]
- Shahali S, Mortimer ST, McLachlan R, O'Bryan MK, Fox DZ, Mortimer D. Refined trajectory smoothing and deep learning classification of human sperm motility. Hum. Reprod 2026(00):1-11. [Google Scholar]
- Li J, Song Y, Dong Y, Guo Q, Wang H, Li Y. Effects of TLR7/8 activator on sperm quality, energy metabolism and the separation efficiency of X- and Y-chromosome-bearing sperm in frozen-thawed bull semen. Indian J. Anim. Res 2025;59(12):2054-2060. [Google Scholar]
- Kotzev K, Metodiev N, Abadjieva D. Application of R848 ligand to improve the efficiency of sex separation in ram spermatozoa. Comptes Rendus Acad. Bulg. Sci 2026;79(1). [Google Scholar]
- Meel M, Gaur M, Punetha M, Dahiya P, Mangal M, Bala R. Use of R848, a TLR7/8 agonist to separate X and Y spermatozoa in buffalo: Effects on cleavage rates, blastocyst production, and the ratio of male to female embryos. Theriogenology 2025;247:117576. [Google Scholar]