


ABSTRACT
Background and Aim: The tropical eel Anguilla bicolor bicolor is a valuable aquaculture commodity; however, the survival of glass eels during the early nursery stage remains low due to nutritional limitations and vulnerability to viral infections, especially Anguillid herpesvirus-1 (AngHV-1). Using microalgae to enrich live feed has been suggested as a strategy to enhance larval nutrition and resilience in endemic disease conditions. This study examined the effects of Artemia enriched with the red microalgae Porphyridium cruentum on the nutritional quality of live feed, survival and growth of glass eels, intestinal structure, water quality, bacterial dynamics, and AngHV-1 infection status under hatchery-like conditions.
Materials and Methods: A completely randomized design with seven treatments was employed: an unenriched control and six enrichment combinations based on P. cruentum cell density and enrichment duration (3 × 105 cells/mL for 6 h; 3 × 105 cells/mL for 12 h; 6 × 105 cells/mL for 6 h; 6 × 105 cells/mL for 12 h; 9 × 105 cells/mL for 6 h; and 9 × 105 cells/mL for 12 h), each with three replicates. Glass eels (0.12 ± 0.01 g; 4.96 ± 0.09 cm) were stocked at 90 fish per tank in 70-L containers and reared for 60 days, fed exclusively with either enriched or unenriched Artemia. Evaluations included the proximate composition of enriched Artemia, survival rate, growth performance, intestinal villus morphometry, water quality parameters, total heterotrophic bacterial counts, and the presence of AngHV-1 verified through conventional polymerase chain reaction.
Results: Enrichment substantially enhanced the nutritional profile of Artemia, increasing crude protein to 61.48% and lipid content to 9.98% on a dry matter basis. The treatment with P. cruentum at 6 × 105 cells/mL for 6 h yielded the highest survival rate (60.74 ± 3.9%) and specific growth rate (1.11 ± 0.1%/day), both significantly higher than those in the control and other treatments (p < 0.05). Intestinal villus length and surface area showed no significant differences among treatments (p > 0.05). Water quality remained within suitable ranges for glass eel culture, and bacterial counts, although slightly higher in enriched treatments, stayed below harmful thresholds. AngHV-1 was detected in all groups, but the enriched treatment at 6 × 105 cells/mL for 6 h exhibited the faintest viral DNA band, indicating a reduction in apparent viral signal.
Conclusion: Short-term enrichment of Artemia with P. cruentum, especially at 6 × 105 cells/mL for 6 h, notably improves the survival and growth performance of A. bicolor bicolor glass eels without harming intestinal morphology or water quality. The lower AngHV-1 polymerase chain reaction signal indicates a possible role of nutritionally enriched live feed in boosting host resilience and reducing viral activity under endemic infection conditions. This enrichment method offers a practical and non-drug approach to enhance early-stage eel aquaculture performance.
Keywords: Anguilla bicolor bicolor, Anguillid herpesvirus-1, Artemia enrichment, eel aquaculture, glass eel survival, Porphyridium cruentum, tropical eel culture, viral resilience.
INTRODUCTION
Tropical eel aquaculture encounters a major hurdle at the glass eel stage, where high and unpredictable mortality rates severely hinder production efficiency and industry growth [1–4]. In Indonesia and other producing countries, A. bicolor bicolor glass eels are still entirely collected from the wild because reliable artificial breeding methods have not yet been developed [5, 6]. Wild-caught glass eels often carry viral and bacterial pathogens acquired during their marine and estuarine life stages, making them highly susceptible to disease outbreaks when transferred to intensive farming systems [1–3]. Among these pathogens, Anguillid herpesvirus-1 (AngHV-1) has been repeatedly identified as a primary cause of mortality in both European and tropical eel species, with high prevalence in wild and farmed populations [1–4]. The reliance on infected wild seed and the lack of effective antiviral treatments make AngHV-1 a significant obstacle to sustainable eel aquaculture [1–4].
Herpesviruses establish lifelong latency in their hosts and can reactivate under stress or immunosuppression, leading to recurrent disease episodes rather than complete clearance [1, 3]. In the case of AngHV-1, eradication from wild glass eel stocks is not feasible with current tools, and direct antiviral drugs are not available for aquaculture use [1–4]. Consequently, disease management in eel hatcheries and nurseries must rely on non-pharmacological strategies that enhance host resilience, limit viral replication, and keep infections in a subclinical or latent state [1, 3]. Previous studies have mainly focused on pathogen detection, epidemiology, or outbreak description, with relatively few efforts to explore nutritional or functional feed approaches as tools to modulate AngHV-1 infection dynamics under endemic conditions [1–4, 7].
Adequate nutrition is essential for the survival and growth of glass eels during the transition from yolk dependence to external feeding [8, 9]. In early life stages, fish larvae and glass eels have high metabolic rates but limited digestive capacity; therefore, they rely on live feeds with suitable particle size, digestibility, and nutrient content [5, 8–10]. Artemia nauplii are still the most commonly used live feed in marine and brackishwater fish culture, including eel nurseries, but their natural levels of long-chain polyunsaturated fatty acids (LC-PUFA), vitamins, and antioxidants are often insufficient [5, 6, 10]. To address these issues, enriching Artemia with microalgae or commercial emulsions has become standard practice, especially to boost n-3 highly unsaturated fatty acids (HUFA), such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which are vital for growth, neural development, and stress resistance in fish larvae [5, 6, 10, 11]. However, most research on Artemia enrichment has concentrated on marine finfish and crustaceans, focusing on growth and survival in pathogen-free settings, and rarely considering viral disease scenarios or glass eels [5, 6, 10, 11].
The red microalgae Porphyridium cruentum is increasingly seen as a promising functional ingredient for aquaculture because of its unique biochemical profile [8, 11]. Besides its relatively high protein and lipid contents, P. cruentum produces large amounts of PUFA (including EPA), phycobiliproteins (such as phycoerythrin), carotenoids, and sulfated polysaccharides with antioxidant, immunomodulatory, and antiviral properties in various aquatic and mammalian models [8, 11]. These qualities indicate that P. cruentum could serve not only as a nutritional supplement for live feeds but also as a functional agent that can influence host–pathogen interactions and boost resistance against viral infections [8, 11]. However, the use of P. cruentum in live-feed enrichment for eel culture is still largely unexplored, and its potential to influence AngHV-1 infection has not been proven through experiments [6, 11].
Furthermore, most studies on eel viral diseases have used challenge models or focused on relatively pathogen-free populations, while commercial hatcheries and nurseries usually face endemic infection pressure, with glass eels already AngHV-1 positive at stocking [1–3]. This discrepancy raises questions about the relevance of traditional challenge trials to real industry conditions [1, 3]. Therefore, there is a need for experimental approaches that simulate actual hatchery scenarios, where the goal is not to prevent initial infection but to suppress viral replication and lessen clinical impacts in already infected glass eel groups [1, 3]. In this context, nutritional strategies that combine better growth performance with partial viral suppression, without relying on drugs or vaccines, could be a crucial step toward creating more biosecure and sustainable fish aquaculture systems [1, 3, 9].
Despite the growing recognition of viral diseases as major challenges in eel aquaculture, most studies on AngHV-1 have mainly concentrated on virus detection, epidemiology, and pathological features rather than on practical mitigation strategies suitable for hatcheries. In particular, the early nursery stage of A. bicolor bicolor remains poorly studied from a nutritional disease-management perspective, even though this stage is linked to the highest mortality rates in commercial production systems. Previous research on Artemia enrichment has mostly targeted marine finfish and crustacean larvae in pathogen-free experimental conditions, primarily aiming to improve growth and survival by enhancing essential fatty acids such as EPA and DHA. However, there is very limited information on how enriched live feeds affect host resilience against viral infections in eel culture, especially under endemic infection scenarios where glass eels already carry AngHV-1 at stocking. Additionally, although P. cruentum is known to contain bioactive compounds like PUFA, phycobiliproteins, carotenoids, and sulfated polysaccharides with antioxidant and immunomodulatory properties, its use as a functional enrichment agent for live feed in eel aquaculture has received little scientific attention. To date, no comprehensive study has simultaneously assessed the nutritional enhancement of Artemia, the growth and survival of glass eels, intestinal histological responses, microbial dynamics, water quality stability, and AngHV-1 infection status under hatchery-like conditions. This significant lack of integrated data leaves an important knowledge gap, especially for hatcheries relying on wild-caught glass eels already exposed to viral pathogens.
Therefore, the current study aimed to assess the effectiveness of Artemia enriched with P. cruentum as a functional live feed for enhancing the performance of A. bicolor bicolor glass eels under conditions where AngHV-1 is endemic. Specifically, this research examined how different combinations of P. cruentum cell density and enrichment duration affected several key biological and environmental factors relevant to early eel cultivation. These included: (i) the proximate composition and estimated energy value of enriched Artemia; (ii) the survival rate and growth performance of glass eels during the nursery stage; (iii) intestinal villus morphology as an indicator of digestive capacity; (iv) water quality parameters and bacterial activity in the rearing environment; and (v) the detection and relative severity of AngHV-1 infection via polymerase chain reaction. The results are expected to offer practical insights for developing nutritionally based management strategies that support sustainable tropical eel aquaculture production.
MATERIALS AND METHODS
Ethical approval
All experimental procedures involving Anguilla bicolor bicolor were conducted in accordance with the institutional guidelines for the care and use of aquatic animals in research and were reviewed and approved by the Ethical Committee of the National Research and Innovation Agency (BRIN), Indonesia, under approval number 253/KE.02/SK/11/2024. The approved protocol covered all stages of the study, including the collection, transportation, acclimatization, stocking, rearing, feeding, handling, sampling, and euthanasia of glass eels used in the experiment.
Particular attention was given to minimizing animal stress and unnecessary suffering throughout the study. Glass eels collected from the Cimandiri River estuary were transported in oxygenated double-layer plastic bags placed in insulated containers with ice gel packs to maintain suitable conditions during transit. On arrival at the experimental facility, the fish were acclimatized for 7 days before the feeding trial to allow recovery from transport-related stress. During the 60-day rearing period, fish were maintained under controlled environmental conditions with continuous aeration and water quality kept within acceptable ranges for eel culture. Handling during routine management and sampling was performed carefully and only when necessary to reduce physical disturbance and stress.
At the end of the experiment, fish sampling for growth assessment, histological examination, and molecular detection of AngHV-1 was carried out in accordance with the approved ethical protocol. The number of fish sampled was kept to the minimum required to achieve the scientific objectives while ensuring reliable data collection. Thus, the study complied with recognized animal welfare principles for the humane use of aquatic organisms in experimental research.
Study period and location
The experiment was conducted over a 5-month period from December 2024 to April 2025 at the Brackishwater Aquaculture Development Center (BADC), Jepara Subdistrict, Jepara Regency, Central Java Province, Indonesia (59418).
Source of P. cruentum and glass eels
The inoculum of P. cruentum was obtained from pure stock cultures maintained at BADC, Jepara, during the intermediate (exponential) growth phase in 20 L glass carboys to ensure culture purity and sufficient cell density. Cultures were kept in f/2 medium [12] at 30 g/L (approximately 30 ppt) salinity and 24°C ± 1°C, under a 12 h light: 12 h dark photoperiod with continuous aeration.
Glass eels of A. bicolor bicolor were collected from local fishers operating in the Cimandiri River estuary, Pelabuhan Ratu Subdistrict, Sukabumi Regency, West Java Province, Indonesia. For transportation, the glass eels were packed in double-layer plastic bags containing 3 L of water with a biomass density of approximately 200 g per bag. The bags were oxygenated and placed in insulated foam boxes containing four ice gel packs (500 g; 20 × 15 × 3 cm). No water exchange was conducted during transport. Upon arrival at the experimental facility, the glass eels were acclimatized for 7 days in 0.5 m³ fiberglass tanks to recover from transport stress and were fed Artemia ad libitum.
Experimental tanks and rearing conditions
After acclimation, glass eels were randomly placed into 70 L plastic containers (53.5 × 37.5 × 32.0 cm³) with a water volume of 30 L. The containers were situated in a semi-indoor facility (200 m²) under a 12 h light and 12 h dark photoperiod. Each container had a single aeration point connected to a root blower system.
Water used for the culture system was sourced from a borewell and allowed to settle for 3 days in a 10 m³ aerated concrete tank. The water was then supplied to the experimental tanks using a 30 W submersible pump through dual cartridge filtration units (Kinnoyama, Nanotech @ SG Clear). The initial mean body weight and total length of the glass eels were 0.12 ± 0.01 g and 4.96 ± 0.09 cm, respectively. Fish were stocked at a density of 90 individuals per tank and reared for 60 days under the respective feeding treatments.
Hatching and enrichment of Artemia with P. cruentum
Artemia cysts (Supreme Plus, Golden West®, Great Salt Lake, USA) were used as live feed. Cysts were hatched following standard procedures commonly used for commercial Artemia production in marine fish larviculture. Briefly, cysts were incubated in conical hatching tanks containing artificial seawater at 30 g/L salinity and 28°C ± 1°C with continuous aeration and illumination for 24 h to obtain instar-I nauplii [13, 14].
For enrichment, newly hatched nauplii were stocked at approximately 300–500 individuals/L in 5 L cylindrical-conical vessels containing seawater at 30 g/L salinity and 28°C ± 1°C. Strong aeration was provided to maintain suspension of both Artemia and microalgal cells. The experimental design consisted of one control treatment (unenriched Artemia) and six enrichment treatments with different combinations of P. cruentum cell density and enrichment duration: control (no enrichment), A (3 × 105 cells/mL for 6 h), B (3 × 105 cells/mL for 12 h), C (6 × 105 cells/mL for 6 h), D (6 × 105 cells/mL for 12 h), E (9 × 105 cells/mL for 6 h), and F (9 × 105 cells/mL for 12 h). Each treatment was replicated three times (n = 3 tanks). Throughout the experimental period, glass eels were fed exclusively with Artemia corresponding to the assigned enrichment treatment.
Growth performance and parameter measurements
At the start of the experiment, 50 fish were randomly sampled to determine initial body weight and total length. At the end of the 60-day trial, all fish from each tank were individually measured to assess final body weight and total length. The growth parameters evaluated included survival rate (SR) [16], average weight gain (AWG) [17], and specific growth rate (SGR).
SR (%) was calculated as: SR = 100 × (Nf/Ni), where Nf represents the final number of fish and Ni represents the initial number of fish.
AWG (g) was calculated as: AWG = Wf − Wi, where Wf and Wi represent the final and initial mean body weights, respectively.
SGR (%/day) was calculated as: SGR = 100 × (ln Wf − ln Wi)/t, where t represents the rearing period (days).
Proximate composition analysis of Artemia
The proximate composition of Artemia (moisture, crude protein, crude lipid, and ash) after enrichment with P. cruentum was determined using standard analytical methods [18–20]. For each treatment, three independent Artemia samples of approximately 30 g wet weight were collected at the end of the enrichment period. The samples were rinsed with distilled water, blotted dry, frozen at −20°C, freeze-dried, and homogenized using a mortar and pestle before analysis [21].
Moisture content was determined according to SNI 01-2891-1992. Crude protein was analyzed using the 18-8-31/MU titrimetric method, whereas crude lipid content was determined using method 18-8-5/MU (Weibull gravimetric method). Ash content was measured following SNI 01-2891-1992 section 6.1. Nitrogen-free extract (NFE, %) was calculated as: NFE = 100 − (moisture + protein + lipid + ash) [22]. Gross energy (kcal/100 g) was estimated using physiological fuel values of 4.0 kcal/g for protein and NFE and 9.0 kcal/g for lipid according to Atwater conversion factors [23, 24].
Intestinal villi morphometric analysis
At the end of the experiment, one glass eel from each replicate tank was sampled for histological examination. Tissue samples were fixed in Bouin’s solution for 48 h and then dehydrated through graded ethanol concentrations (70%, 80%, 90%, 95%, and 100%), followed by immersion in an ethanol–xylol solution (1:1). The tissues were embedded in paraffin blocks and sectioned transversely at the abdominal region (from the posterior part of the stomach to the anus) using a microtome. Histological sections were stained with hematoxylin–eosin.
Microscopic observations were performed using an Olympus BX53 microscope (Japan) at 10× magnification. Villus length and surface area were calculated with the modified formula: villus surface area (µm²) = [(b + c)/2] × a, where a represents villus height or length, b represents apical villus width, and c represents basal villus width.
Detection of AngHV-1 infection
Detection of AngHV-1 infection in the A. bicolor bicolor glass eel population was carried out twice: once before the experiment began and again at the end of the rearing period. Viral genome detection was performed qualitatively (semi-quantitatively) using conventional polymerase chain reaction based on the method described previously [25], with minor modifications.
For each treatment and sampling time, gill and kidney tissues from five glass eels (one fish from each replicate tank) were pooled to create a single DNA sample. Genomic DNA was extracted using a commercial tissue DNA extraction kit (Geneaid™ DNA Isolation Kit Ver. 06.22.16) following the manufacturer’s instructions. The quality and concentration of the extracted DNA were measured with a NanoDrop 2000 UV-Vis spectrophotometer (Thermo Scientific, WA, USA).
Detection of AngHV-1 was carried out using the primer pair HVAPOLVPSD and HVAPOLOOSN (Table 1), which amplify a 394-bp fragment of the viral genome [25]. PCR amplification was performed under the following conditions: initial denaturation at 94°C for 5 min (1 cycle), followed by 40 cycles of denaturation at 94°C for 30 s, primer annealing at 65°C for 45 s, and extension at 72°C for 60 s, with a final extension step at 72°C for 7 min. PCR products were stored at 4°C until analysis and subsequently visualized using 2% agarose gel electrophoresis to confirm the presence of viral DNA bands.
Table 1. Primers used for detection of Anguillid herpesvirus 1.
| Primary name | Primary sequence | Reference |
|---|---|---|
| HVAPOLVPSD | 5′-GTG TCG GGC TTT GTG GTG C-3′ | [25] |
| HVAPOLOOSN | 5′-CAT GCC GGG AGT CTT TTT GAT-3′ | [25] |
Statistical analysis
All results are presented as mean ± standard error (SE), with the tank considered as the experimental unit. Before conducting statistical analysis, the data were tested for normality using the Shapiro–Wilk test and for homogeneity of variance with Levene’s test. When these assumptions were met, a one-way analysis of variance was used to assess the effect of treatments. If significant differences were found (p < 0.05), Duncan’s multiple range test was applied as a post hoc test to compare treatment means. Statistical analyses were conducted using SPSS software version 31 (IBM Corp., NY, USA).
RESULTS AND DISCUSSION
Nutrient composition of enriched Artemia
The proximate composition and energy content of enriched Artemia are shown in Table 2. Moisture content, expressed on a wet weight basis, ranged from 91.83% to 94.21%, while crude protein, crude lipid, ash, NFE, and gross energy were expressed on a dry matter basis. Compared to the control, enrichment consistently enhanced the nutritional value of Artemia. Crude protein increased from 47.72% in the control to 48.28%–61.48% in the enriched treatments, with the highest level observed in treatment A. Even greater increases were seen in crude lipid, which rose from <0.30% in the control to 4.72%–9.98% in the enriched groups, reaching a maximum in treatment F. This rise in lipid content contributed to higher fat-based energy, which increased from a negligible amount in the control to 3.29–5.67 kcal/100 g in the enriched treatments.
Total gross energy increased across all enriched groups, with the highest values seen in treatments C (26.74 kcal/100 g) and E (26.71 kcal/100 g), compared to 18.90 kcal/100 g in the control. Conversely, ash and NFE showed variable patterns among treatments. These findings suggest that enrichment with P. cruentum enhanced not only the macronutrient profile of Artemia but also its overall energy value.
The significant increase in crude protein and crude lipid highlights P. cruentum’s effectiveness as a bioencapsulation agent. Artemia’s active filtration behavior helps it absorb biomolecules from P. cruentum, including proteins, unsaturated fatty acids, and lipophilic pigments [26]. The biochemical makeup of P. cruentum includes protein (30%–43%), lipid (9%–14%), and EPA, which makes up about 9.7% of total fatty acids [27]. Additionally, compounds like sulfated polysaccharides and phycoerythrin further boost the nutritional and biological value of live feed [28, 29]. Therefore, enrichment with P. cruentum enhances both the quantity and quality of Artemia.
Among the tested treatments, C offered the most balanced nutrient profile, with crude protein at 52.59%, crude lipid at 4.72%, and total energy of 26.74 kcal/100 g. The 6 h enrichment period likely coincided with the peak ingestion phase of Artemia on microalgal cells [26], making it an efficient enrichment window [30]. The increased energy from lipid and pigment fractions, along with the antioxidant and bioactive properties of P. cruentum, may also enhance the immunoprotective value of enriched Artemia [27, 31]. Overall, these findings suggest that enrichment with P. cruentum improves Artemia’s nutritional profile and enhances its suitability as a strategic live feed for the early rearing phase of A. bicolor bicolor.
Table 2. Proximate composition of Artemia nauplii: moisture (% wet weight), crude protein, crude lipid, ash, nitrogen-free extract (NFE) (% dry matter), and gross energy (kcal/100 g dry matter) in control and Porphyridium cruentum-enriched treatments (A–F).
| Parameter | Control | A | B | C | D | E | F |
|---|---|---|---|---|---|---|---|
| Moisture (%) | 93.42 | 93.60 | 94.04 | 92.27 | 94.21 | 91.83 | 93.69 |
| Crude protein (%) | 47.72 | 61.48 | 48.28 | 52.59 | 49.14 | 51.68 | 51.26 |
| Crude lipid content (%) | <0.30 | 6.33 | 7.88 | 4.72 | 6.21 | 5.69 | 9.98 |
| Ash (%) | 28.19 | 29.06 | 27.66 | 19.54 | 32.59 | 19.08 | 21.38 |
| NFE* (%) | 23.80 | 3.13 | 16.18 | 23.15 | 12.06 | 23.55 | 17.38 |
| Total energy (kcal/100 g) | 18.90 | 20.10 | 19.59 | 26.74 | 17.31 | 26.71 | 23.01 |
Survival rate
The survival of A. bicolor bicolor over the 60-day rearing period varied significantly among treatments (Figure 1). The control group had the lowest survival rate at 15.56% ± 5.9%, while treatment C achieved the highest survival at 60.74% ± 3.9%, which was significantly higher than all other groups (p < 0.05). Treatments A (28.89% ± 9.2%), B (20.00% ± 9.5%), D (27.41% ± 9.3%), and F (26.67% ± 6.9%) did not differ significantly from the control (p > 0.05). Treatment E (32.78% ± 2.4%) showed significantly higher survival than the control but remained lower than treatment C. These findings demonstrate that enriching Artemia with P. cruentum at 6 × 105 cells/mL for 6 h was the most effective condition for improving survival during rearing.
The improved survival rate is likely due to the increased supply of nutrients and bioactive compounds transferred from P. cruentum to Artemia. Higher lipid and protein levels provide vital energy and substrates for larval development [32, 33]. Lipids act as a key energy reserve during metamorphosis, while proteins support tissue repair and growth. Additionally, PUFA, especially DHA, along with phycoerythrin from P. cruentum, may enhance energy metabolism, enzymatic activity, membrane fluidity, and osmoregulation [33, 34]. Functional compounds such as sulfated polysaccharides and EPA may also boost larval immunity and thus improve survival [28, 35].
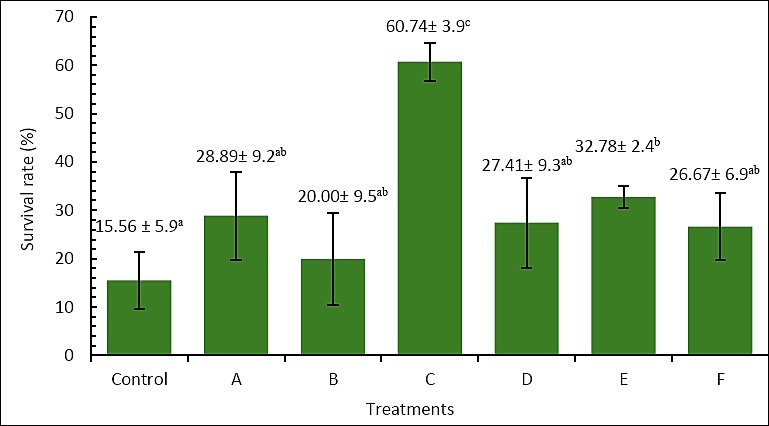
Figure 1. Survival (%) of Anguilla bicolor bicolor glass eels after 60 days of rearing under different combinations of Artemia enrichment density and duration using Porphyridium cruentum. Remark: Values followed by the same superscript letter are not significantly different (p > 0.05).
Growth performance
Final body weight after 60 days also varied significantly among treatments (Figure 2). The control group had the lowest final body weight, at 0.181 ± 0.02 g, while treatment C showed the highest value, at 0.250 ± 0.01 g, which was significantly higher than all other treatments (p < 0.05). Treatments A (0.197 ± 0.03 g), B (0.199 ± 0.03 g), D (0.203 ± 0.02 g), E (0.191 ± 0.02 g), and F (0.186 ± 0.02 g) did not differ significantly from the control (p > 0.05). These results suggest that treatment C was also the most effective in increasing final body weight.
A similar pattern was observed for SGR (Figure 3). Treatment C yielded the highest SGR, at 1.11 ± 0.1%/day, which was significantly greater than the control (0.65 ± 0.1%/day) and all other treatments (p < 0.05). In contrast, treatments A (0.69 ± 0.2%/day), B (0.59 ± 0.6%/day), D (0.82 ± 0.1%/day), E (0.84 ± 0.1%/day), and F (0.80 ± 0.2%/day) did not differ significantly from the control (p > 0.05). Therefore, enriching Artemia with P. cruentum at 6 × 105 cells/mL for 6 h produced the greatest improvement in both final body weight and SGR of A. bicolor bicolor.
These growth responses align with the nutrient profile shown in Table 2. Treatment C offered a balanced mix of crude protein, crude lipid, and total energy, ensuring an adequate supply of amino acids and energy for larval growth. Protein serves as the main substrate for tissue synthesis and enzyme production, while lipid, especially PUFA, functions as a key energy reserve during metamorphosis and developmental transitions [32, 33]. Since zooplankton naturally provide a major lipid source for larvae, enriching Artemia with P. cruentum may better mimic natural feeding conditions by increasing the availability of long-chain fatty acids essential for larval nutrition [33, 34].
The physiological value of P. cruentum is further supported by its high content of PUFA, including EPA and DHA precursors, as well as phycoerythrin and sulfated polysaccharides. These compounds may boost membrane fluidity, metabolic efficiency, osmoregulation, and immune responses [28, 34, 35]. Transferring these compounds into Artemia enhances the functional quality of the live feed by providing not just energy but also biologically active substances that aid larval physiology and resilience [36–38]. The simultaneous increase in survival and growth suggests that nutritional improvements benefited both parameters at the same time. Since these gains were not linked to changes in intestinal villus morphology, the improved performance in treatment C seems to be mainly due to better nutrient availability and bioactive quality rather than structural modifications of the digestive tract.
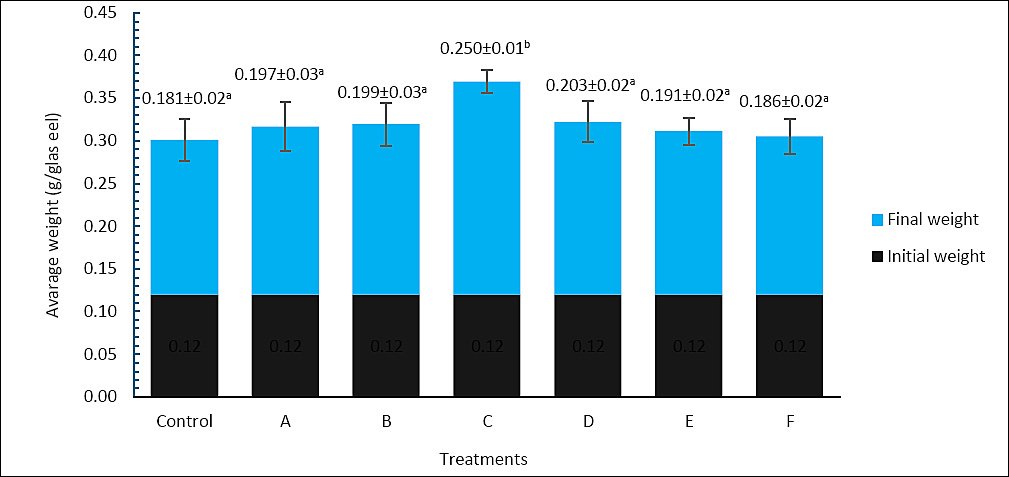
Figure 2. Final body weight (g) of Anguilla bicolor bicolor glass eels after 60 days of rearing under different combinations of Artemia enrichment density and duration using Porphyridium cruentum. Values followed by the same superscript letter are not significantly different (p > 0.05).
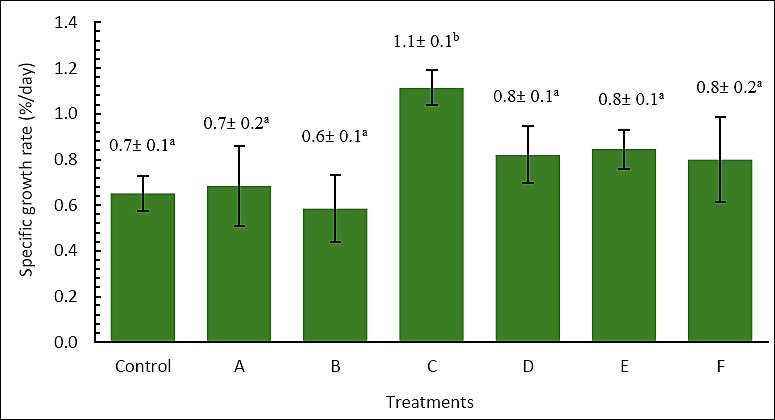
Figure 3. Specific growth rate (%/day) of Anguilla bicolor bicolor glass eels after 60 days of rearing under different combinations of Artemia enrichment density and duration using Porphyridium cruentum. Values followed by the same superscript letter are not significantly different (p > 0.05).
Intestinal villi morphometric analysis
Histological examination of the intestine at the end of the rearing period mostly showed uniform villus structure across treatments (Figure 4). No significant differences were seen between the control and enriched groups. This was supported by morphometric analysis, where villus length ranged from 95.00 to 147.06 µm and villus surface area ranged from 3,717.25 to 6,527.76 µm², with no significant differences among treatments (p > 0.05) (Figure 5). These results suggest that enrichment with P. cruentum, regardless of cell density or duration, did not change intestinal histology, and the basic digestive structure remained similar across treatments.
Intestinal villi are finger-like projections that enhance the surface area for absorption, thereby aiding in the uptake of nutrients and water. Villus height and surface area are frequently used as markers of nutrient absorption ability [39]. Increased villus height, fold number, and surface area generally correlate with better nutrient absorption and utilization because a larger epithelial surface is available for diffusion and active transport [40].
In the present study, although enrichment with P. cruentum significantly enhanced the growth performance of A. bicolor bicolor, no corresponding changes were observed in villus morphometry. Villus length and surface area remained relatively unchanged across treatments, indicating that dietary enrichment did not cause histological adaptation in the intestine. Therefore, the increased growth seen in treatment C cannot be attributed to an expanded intestinal structural capacity.
These results indicate that P. cruentum mainly contributed by increasing metabolic energy availability rather than altering intestinal structure. Red microalgae are rich in polysaccharides, proteins, and functional lipids such as PUFA, which can boost energy density and metabolic efficiency in larvae [22, 34]. Therefore, the extra energy supplied through enriched Artemia was more likely used for somatic growth and metabolic activities without changing villus morphology. In summary, P. cruentum enrichment seems to support the growth of A. bicolor bicolor by improving nutrient and bioactive compound availability while keeping the intestinal structure stable.
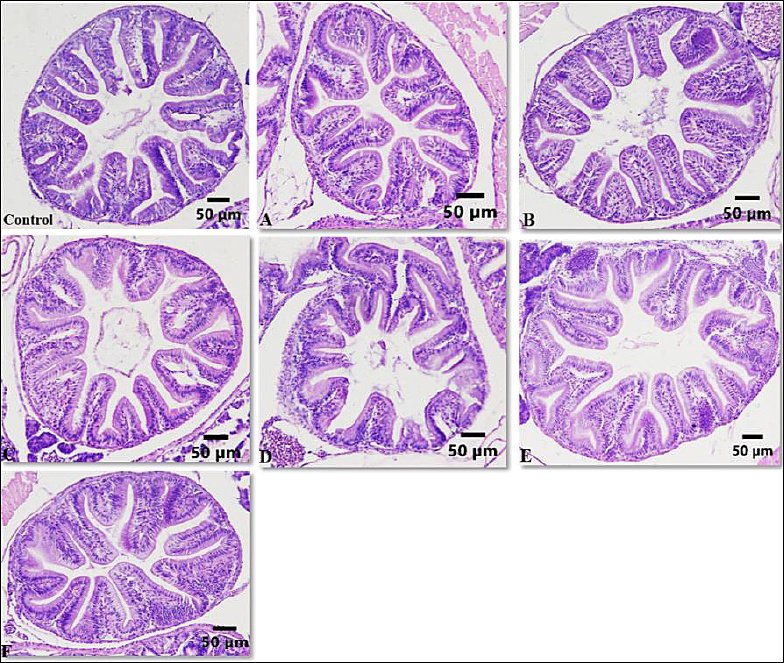
Figure 4. Histological cross-sections of the intestine of Anguilla bicolor bicolor glass eels fed Artemia enriched with Porphyridium cruentum at varying densities and enrichment durations.
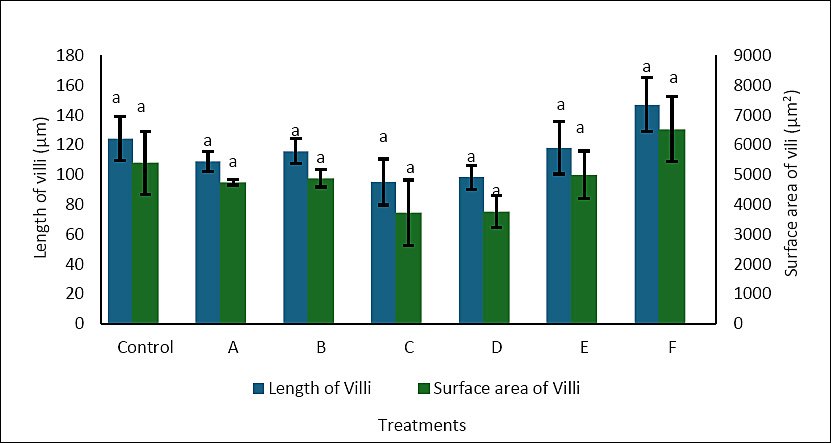
Figure 5. Villus length (μm) and villus surface area (μm²) in the intestine of Anguilla bicolor bicolor glass eels following feeding with Artemia enriched with Porphyridium cruentum under different density and enrichment time combinations.
Water quality parameters
Water quality remained fairly constant across treatments during the rearing of A. bicolor bicolor (Table 3). DO ranged from 4.60 to 6.50 mg/L, temperature from 26.80°C to 28.00°C, pH from 7.26 to 8.55, and total ammonia nitrogen (TAN) from 0.001 to 0.448 mg/L. These values stay within acceptable ranges for the growth and development of A. bicolor bicolor and suggest that no significant environmental fluctuations took place during the culture period. This stability also indicates that using Artemia enriched with P. cruentum did not negatively impact water quality.
High-quality and digestible live feed like Artemia is known to reduce uneaten feed residues and nitrogenous waste, thereby decreasing the risk of water pollution [41]. In this study, enrichment with P. cruentum not only enhanced the nutritional profile of Artemia, including water-soluble proteins, PUFA, and antioxidants, but also seemed to support environmental stability. Red microalgae are recognized for their ability to absorb nitrogen, which may indirectly help keep TAN within safe limits [42]. Additionally, several compounds in P. cruentum have antioxidant, anti-inflammatory, and immunomodulatory properties that could reduce stress and assist in maintaining physiological homeostasis in cultured fish [43, 44].
This observation aligns with previous reports suggesting that supplementing aquafeeds with microalgal products can enhance not only nutritional quality but also fish health and water quality stability [45, 46]. Therefore, maintaining suitable water quality in this study probably reflects a positive interaction between the improved nutritional performance of enriched Artemia and the ecological contribution of P. cruentum in the rearing system.
Table 3. Ranges of water quality parameters (dissolved oxygen [DO], temperature, pH, and total ammonia nitrogen [TAN]) recorded during the 60-day rearing period of Anguilla bicolor bicolor glass eels under different Artemia enrichment treatments.
| Treatment | DO (mg/L) | Temperature (°C) | pH | TAN (mg/L) |
|---|---|---|---|---|
| Control | 4.6–6.4 | 26.8–28.0 | 7.26–8.18 | 0.275–0.428 |
| A | 4.6–6.3 | 26.8–27.8 | 7.91–8.48 | 0.001–0.133 |
| B | 4.6–6.5 | 26.8–28.0 | 7.92–8.45 | 0.120–0.164 |
| C | 4.6–6.5 | 26.8–27.9 | 7.94–8.48 | 0.276–0.448 |
| D | 4.6–6.5 | 26.8–27.9 | 7.93–8.49 | 0.076–0.394 |
| E | 4.6–6.4 | 26.8–27.8 | 7.93–8.55 | 0.040–0.253 |
| F | 4.7–6.4 | 26.9–27.9 | 7.93–8.40 | 0.273–0.403 |
Bacterial dynamics in the rearing medium
Although water quality stayed within acceptable limits, microbiological conditions also significantly influenced the culture environment. Total plate count (TPC) analysis revealed that bacterial counts were higher in all enriched treatments compared to the control (0.84 × 105 CFU/mL), with the highest count found in treatment D (2.00 × 105 CFU/mL). The other enriched groups ranged from 1.00 × 105 to 1.80 × 105 CFU/mL (Figure 6). Despite this increase, all counts remained within levels typically regarded as safe for cultured fish.
The higher bacterial counts in treatments receiving P. cruentum-enriched Artemia may be attributed to two factors. First, enriched Artemia might have carried additional biomolecules, such as water-soluble proteins, sulfated polysaccharides, and lipophilic pigments, which can serve as substrates for heterotrophic bacteria [36, 38]. Second, red microalgae like P. cruentum are known to produce EPS [35], which may act as carbon sources for microbial communities while also providing immunostimulatory effects in fish.
Moderate bacterial abundance may not necessarily be harmful. Non-pathogenic bacterial communities at low to moderate levels can prevent colonization by pathogenic bacteria through competition for nutrients and space, while also producing antagonistic bioactive metabolites [47]. Therefore, the increase in TPC observed in enriched treatments should not be seen only as a negative result but may instead reflect a more active and potentially beneficial microbial environment.
Importantly, the increased bacterial load did not harm growth performance or survival. This indicates that the beneficial nutritional and bioactive properties of P. cruentum outweighed any potential risks linked to higher bacterial levels. Therefore, enrichment with P. cruentum not only enhanced the nutritional quality of Artemia but may also have affected microbial interactions in the culture system in a way that is compatible with glass eel rearing.
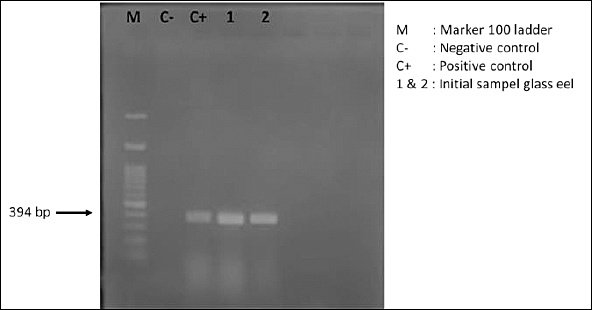
Figure 6. Agarose gel electrophoresis profile showing DNA amplicons of Anguillid herpesvirus 1 in Anguilla bicolor bicolor glass eels prior to their use as experimental test organisms.
Detection of AngHV-1 infection
Molecular screening conducted before the experiment confirmed that the A. bicolor bicolor population used in this study was already infected with AngHV-1 (Figure 6). This finding aligns with previous reports showing that wild-caught glass eels commonly harbor viral infections, which pose a major challenge for aquaculture [48, 49]. Semi-quantitative assessment based on DNA band intensity indicated that infection levels were already high at the start of rearing.
After 60 days, polymerase chain reaction analysis showed that AngHV-1 genomic DNA was still detectable in all experimental groups, although band intensity varied among treatments (Figure 7). The control and treatments A, B, D, E, and F exhibited relatively thick bands, indicating severe infection. Treatment B had a moderate band intensity, while treatment C showed only a faint band, consistent with mild infection. These findings suggest that dietary supplementation through Artemia enriched with P. cruentum may lessen the severity of AngHV-1 infection, although it does not completely eliminate the virus.
This pattern can be understood within the framework of herpesvirus biology. As part of the Herpesviridae family, AngHV-1 establishes lifelong latency, with the viral genome remaining in host cell nuclei as episomes and reactivating under stress or immunosuppression [50]. Therefore, although viral DNA was detectable in all treatments, the fainter bands in treatments C and B indicate lower viral activity and suggest the virus may be in a more latent state. This suggests that improved physiological and immunological health in the enriched groups may have suppressed viral replication.
These results also align with previous reports indicating that AngHV-1 is widespread among various Anguilla species, including A. anguilla, A. japonica, A. rostrata, A. bicolor bicolor, and A. marmorata [48, 49, 51–54]. Since artificial propagation of glass eels is not yet available [55, 56], eel aquaculture still relies on wild-caught seed that may already carry infections before entering culture systems. Given these circumstances, completely preventing infection is unrealistic, and management should focus on minimizing viral impact rather than stopping exposure.
Since herpesviruses are obligate intracellular pathogens, direct chemotherapeutic control is highly limited [48]. Therefore, managing AngHV-1 must depend on non-drug approaches. These can include functional feeds and immunostimulants, vaccination, and environmental adjustments such as temperature control to inhibit viral replication [52]. Elevated temperatures have also been reported to boost antiviral responses by increasing the expression of immune-related genes, including interferon-γ, interleukin-1β, and Mx1 [20]. While these methods may not eradicate the virus, they can decrease viral replication and reduce the risk of transmission.
In the present study, the relationship among survival (Figure 1), growth performance (Figures 2 and 3), and AngHV-1 band intensity (Figures 7 and 8) indicates that enrichment with P. cruentum supports the physiological condition of A. bicolor bicolor in a way that promotes improved performance while reducing apparent viral severity. Collectively, these results suggest that nutritional management through P. cruentum bioencapsulation plays a key role in mitigating infection severity and maintaining the performance of A. bicolor bicolor glass eels during culture, even though AngHV-1 remains detectable in all groups.
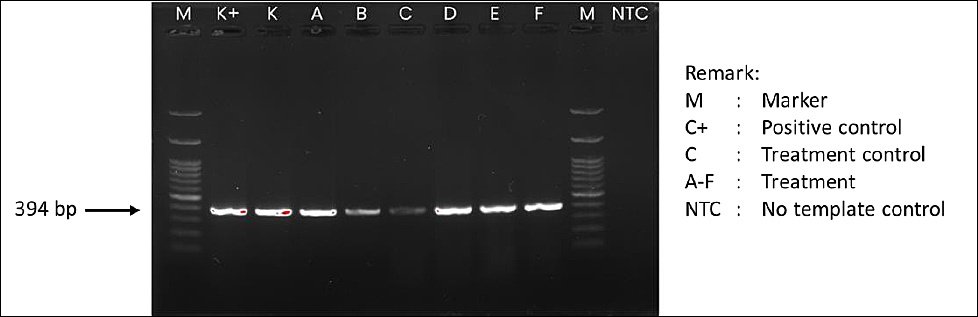
Figure 7. Agarose gel electrophoresis profile showing DNA amplicons of Anguillid herpesvirus 1 in Anguilla bicolor bicolor glass eels after 60 days of rearing under the experimental treatments.
Study limitations
Several limitations should be considered when interpreting the present findings. First, AngHV-1 load was assessed using conventional polymerase chain reaction and semi-quantitative visual evaluation of band intensity, without employing an internal control gene or quantitative PCR-based quantification. Therefore, the differences in AngHV-1 levels among treatments should be regarded as approximate and require confirmation using more sensitive, quantitative molecular methods. Second, intestinal histological assessment was performed on a limited number of fish and villi per treatment, which may have reduced the ability to detect subtle morphometric differences and could explain the lack of significant variation in villus structure despite the notable improvements in survival and growth.
Third, only total heterotrophic bacterial counts were measured, without distinguishing beneficial bacteria from potentially pathogenic taxa. Additionally, water quality was evaluated at specific sampling points rather than continuously, which limited the temporal resolution of microbial and environmental fluctuations during the culture period. Finally, the experiment was conducted at a pilot semi-indoor scale for 60 days with a single cohort of glass eels from one geographic source. Therefore, caution is necessary when extrapolating these findings to other eel stocks, hatchery systems, or long-term production conditions.
Future perspectives
Future research should quantify AngHV-1 dynamics using quantitative PCR and include immune-related endpoints, such as the expression of antiviral and pro-inflammatory genes. It should also incorporate controlled challenge studies to better clarify how P. cruentum-enriched Artemia affects viral replication and host defense. A more comprehensive histological and functional assessment of the digestive tract, including digestive enzyme activity and gut barrier markers, would also help differentiate the relative roles of structural and metabolic factors in improving growth performance.
Additionally, more detailed profiling of crude lipid, fatty acids, and pigments in enriched Artemia, along with characterization of related microbial communities, would offer a more thorough understanding of how P. cruentum alters the nutritional and microbial environment of glass eels. Finally, validation in commercial hatchery settings, including economic analysis and integration with complementary management strategies such as temperature control or vaccination, is necessary to assess the practicality of P. cruentum-enriched Artemia as a non-pharmaceutical approach for managing AngHV-1 in tropical eel aquaculture.
CONCLUSION
This study showed that enriching Artemia with P. cruentum significantly enhances the nutritional quality of live feed and improves the survival and growth of A. bicolor bicolor glass eels under AngHV-1 endemic conditions. Among the tested treatments, enrichment at 6 × 105 cells/mL for 6 h produced the best results, achieving the highest survival rate (60.74% ± 3.9%) and SGR (1.11% ± 0.1%/day), along with a notable boost in Artemia’s nutritional profile, including crude protein, crude lipid, and gross energy content. Despite these notable improvements in survival and growth, intestinal villus morphology remained similar across treatments, suggesting that performance gains were mainly due to enhanced nutrient availability and bioactive compound delivery rather than structural changes in the digestive tract. Additionally, water quality parameters and bacterial levels stayed within acceptable ranges for eel culture, confirming that P. cruentum enrichment did not negatively impact the rearing environment.
From a practical standpoint, these findings emphasize the potential of P. cruentum-enriched Artemia as an effective live feed strategy to improve early rearing success of A. bicolor bicolor in hatchery systems that depend on wild-caught glass eels. The decrease in AngHV-1 DNA band intensity seen in the optimal treatment indicates that better nutritional status and the presence of bioactive compounds from P. cruentum may help increase host resilience and partially suppress viral activity. Since there are no current direct antiviral treatments for AngHV-1, using enriched live feeds for nutritional management offers a practical, non-pharmacological way to reduce disease impacts in eel aquaculture.
A major strength of this study is its comprehensive evaluation of nutritional, physiological, microbiological, and virological factors within a single experimental setup that mimics hatchery conditions. By analyzing feed composition, growth performance, intestinal structure, water quality, bacterial behavior, and AngHV-1 infection status simultaneously, the study offers a thorough understanding of how functional live feed enrichment affects both fish performance and the culture environment.
Overall, the results show that enriching Artemia with P. cruentum at 6 × 105 cells/mL for 6 h is an effective nutritional strategy for boosting survival and growth while enhancing host resilience against AngHV-1 in A. bicolor bicolor glass eels. These findings provide a scientific basis for integrating microalgal enrichment into eel hatchery management and support the development of more sustainable and biosecure tropical eel aquaculture systems.
DATA AVAILABILITY
All data generated or analyzed during this study are included in this manuscript. Additional supporting data are available from the corresponding author upon reasonable request.
AUTHORS’ CONTRIBUTIONS
AS: Conceptualization, data curation, formal analysis, investigation, methodology, and writing of the original draft. LHS: Data curation, formal analysis, project administration, visualization, writing of the original draft, and writing–review and editing. EKMK: Conceptualization, investigation, methodology, and data curation. AN: Investigation, formal analysis, methodology, resources, and writing of the original draft. EBK: Data curation, resources, formal analysis, and writing–review. TT: Investigation, formal analysis, methodology, resources, and writing of the original draft. YRW: Supervision, validation, writing, and resources. RS: Formal analysis, validation, writing of the original draft, and resources. SM: Project administration, methodology, resources, and writing–review and editing. LR: Formal analysis, validation, writing of the original draft, and resources. THP: Data curation, formal analysis, methodology, and supervision. BP: Resources, formal analysis, validation, writing–review and editing, and supervision. TB: Conceptualization, investigation, methodology, data curation, and supervision. MM: Validation, resources, and writing–review and editing. ES: Conceptualization, investigation, methodology, and data curation. HH: Validation, writing–review and editing, and supervision. All authors have read and approved the final manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the financial support from Riset dan Inovasi untuk Indonesia Maju (RIIM) number 7314/III.4.9/HK/12/2023. We also extend our sincere appreciation to the Research Center for Fisheries, Research Organization for Earth Sciences and Maritime BRIN, Mina Sidat Bersatu Cooperative, and IPB University for providing facilities and collaborative assistance throughout this study.
REFERENCES
- Guo R, Zhang Z, He T, Li M, Zhuo Y, Yang X, Chen X. Isolation and identification of a new isolate of Anguillid herpesvirus 1from farmed American eels (Anguilla rostrata) in China. Viruses;2022(14):2722. [Google Scholar] | [Crossref]
- Panicz R, Eljasik P, Nguyen TT, Vo Thi KT, Hoang DV. First detection of Herpesvirus anguillae (AngHV-1) associated with mortalities in farmed giant mottled eel (Anguilla marmorata) in Vietnam. J Fish Dis 2021;44(6):847-52. [Google Scholar] | [Crossref]
- Kullmann B, Adamek M, Steinhagen D, Thiel R. Anthropogenic spreading of Anguillid herpesvirus 1by stocking of infected farmed European eels, Anguilla anguilla (L.), in the Schlei fjord in northern Germany. J Fish Dis 2017;40(11):1695-706. [Google Scholar] | [Crossref]
- Kole S, Kim HJ, Jung SJ. Complete genome sequence of Anguillid herpesvirus 1isolated from imported Anguilla rostrata (American eel) from Canada. Microbiol Resour Announc 2022;11(12):e00829-22. [Google Scholar] | [Crossref]
- Lavens P, Sorgeloos P. Manual on the production and use of live food for aquaculture. Rome: FAO; 1996. p. 295. [Google Scholar]
- Van Stappen G, Sorgeloos P, Rombaut G. Manual on Artemia production and use. Rome: FAO; 2024. [Google Scholar]
- Chen X, Chen H, Ge JQ. Development and application of a recombinase-aided amplification combined with a lateral flow dipstick assay for rapid visual detection of anguillid herpesvirus 1. J Fish Dis 2024;47(4):e13907. [Google Scholar] | [Crossref]
- Li P, Song Z, Huang L, Sun Y, Sun Y, Wang X, Li L. Effects of dietary protein and lipid levels in practical formulation on growth, feed utilization, body composition, and serum biochemical parameters of growing rockfish Sebastes schlegeli. Aquac Nutr 2023;2023(1):9970252. [Google Scholar] | [Crossref]
- Manam VK. Fish feed nutrition and its management in aquaculture. Int J Fish Aquat Stud 2023;11(2):58-61. [Google Scholar] | [Crossref]
- Palma J, Bureau DP, Andrade JP. Effect of different Artemia enrichments and feeding protocol for rearing juvenile long snout seahorse, Hippocampus guttulatus. Aquaculture 2011;318((3-4)):439-43. [Google Scholar] | [Crossref]
- Støttrup J, McEvoy L. Live feeds in marine aquaculture. Berlin: Blackwell Science; 2003. p. 337. [Google Scholar]
- Lananan F, Jusoh A, Ali NA, Lam SS, Endut A. Effect of conway medium and f/2 medium on the growth of six genera of South China Sea marine microalgae. Bioresour Technol 2013;141:75-82. [Google Scholar] | [Crossref]
- Léger P, Bengtson DA, Sorgeloos P, Simpson KL, Beck AD. The nutritional value of Artemia:a review. Wetteren: Universa Press; 1987. p. 556. [Google Scholar]
- Millán-Almaraz MIM, Nieves-Soto MN, López-Peraza DJL, Peraza-Yee MM. Effect of light and feed density on ingestion rate, protein and lipid content of Artemia franciscana juveniles. Lat Am J Aquat Res;2021(49):717-24. [Google Scholar] | [Crossref]
- Montiel TDNJM, Canché LGR. Biomassas production and nutritional value of Artemia sp. (Anostraca:Artemiidae) in Campeche, México. Rev Biol Trop 2005;53((3-4)):447-54. [Google Scholar] | [Crossref]
- Park SW, Jung EB, Kim DW. Outbreak of Anguillid herpesvirus-1 (AngHV-1) infection in cultured shortfin eel (Anguilla bicolor) in Korea. J Fish Pathol 2012;25:151-8. [Google Scholar] | [Crossref]
- Millán-Almaraz MIM, Nieves-Soto MN, López-Peraza DJL, Peraza-Yee MM. Effect of light and feed density on ingestion rate, protein and lipid content of Artemia franciscana juveniles. Lat Am J Aquat Res;2021(49):717-24. [Google Scholar] | [Crossref]
- 01-2891-1992 The water content analysis 1992. [Google Scholar] | [Crossref]
- 18-8-31/MU/SMM-SIG method protein levels were measured through the Titrimetric. [Google Scholar] | [Crossref]
- 18-8-5/MU/SMM-SIG point 3.2.2 total fat content was determined using the Weibull Test (gravimetry). [Google Scholar] | [Crossref]
- Rizk EST, Shoukr FA, El-Gamal MM, Abdel-Razek FA, Mona MM. An attempt to improve the proximate composition of local Artemia strain (Wadi El Natrun, Egypt). J Basic Appl Zool 2018;79(1):24. [Google Scholar] | [Crossref]
- Asgharpour M, Rodgers B, Hestekin JA. Eicosapentaenoic acid from Porphyridium cruentum:Increasing growth and productivity of microalgae for pharmaceutical products. Energies;2015(8):10487-503. [Google Scholar] | [Crossref]
- Webster CD, Lim C. Nutrient requirements and feeding of finfish for aquaculture. Wallingford: CABI; 2002. [Google Scholar]
- Chapter 3:Calculation of the energy content of foods-Energy conversion factors. Report of a Technical Workshop. FAO Food and Nutrition Paper 77. Rome:Food and Agriculture Organization of the United Nations 2003. [Google Scholar] | [Crossref]
- Rijsewijk F, Pritz-Verschuren S, Kerkhoff S, Botter A, Willemsen M, van Nieuwstadt T, Haenen O. Development of polymerase chain reaction for the detection of Anguillid herpesvirus DNA in eel based on the herpesvirus DNA polymerase gene. J Virol Methods 2005;124:8887-94. [Google Scholar] | [Crossref]
- Jiang M, Hu Z, Chen XD, Wu P. Unlocking the release, digestion and absorption kinetics of DHA from different fish oil delivery systems:An in vitro and ex vivo study. Food Hydrocoll 2024;162:110966. [Google Scholar] | [Crossref]
- Van Stappen G. 4.1 Introduction, biology and ecology of Artemia. Rome: FAO; 1996. [Google Scholar]
- Samat NA, Yusoff FM, Rasdi NW, Karim M. Enhancement of live food nutritional status with essential nutrients for improving aquatic animal health:A review. Animals 2020;10(12):1-27. [Google Scholar] | [Crossref]
- El-khodary GM, Mona MM, El-sayed HS, Ghoneim AZ. Phylogenetic identification and assessment of the nutritional value of different diets for a copepod species isolated from Eastern Harbor coastal region. Egypt J Aquat Res;2020(46):173-80. [Google Scholar] | [Crossref]
- Grayson JD, Dabrowski K. Utilization of live-food enrichment with polyunsaturated fatty acids (pufa) for the intensive culture of yellow perch larvae. N Am J Aquac 2022;84(2):131-48. [Google Scholar] | [Crossref]
- Tounsi L, Hlima HB, Hentati F, Hentati O, Derbel H, Michaud P, Abdelkafi S. Microalgae:A promising source of bioactive phycobiliproteins. Mar Drugs 2023;21(8):440. [Google Scholar] | [Crossref]
- Naiel MAS, Negm SS, Ghazanfar S, Shokri M, Abdelnour SA. The risk assessment of high-fat diet in farmed fish and its mitigation approaches:A review. J Anim Physiol Anim Nutr 2023;107(3):948-69. [Google Scholar] | [Crossref]
- Begho T, Irabor AE. Fish feed formulation:Does Nigerian farmers'risk and time preference play a part in choosing feed protein sources for intensively farmed fish?. Aquaculture 2024;585:740723. [Google Scholar] | [Crossref]
- Gendron TF, Bieniek KF, Zhang YJ. Antisense transcripts of the expanded C9ORF72 hexanucleotide repeat form nuclear RNA foci and undergo repeat-associated non-ATG translation in c9FTD/ALS. Acta Neuropathol 2013;126:829-44. [Google Scholar] | [Crossref]
- Su G, Jiao K, Chang J, Li Z, Guo X, Sun Y, Zeng X, Lu Y, Lin L. Enhancing total fatty acids and arachidonic acid production by the red microalgaee Porphyridium purpureum. Bioresour Bioprocess 2016;3((1)). [Google Scholar] | [Crossref]
- Safi C, Charton M, Pignolet O, Pontalier PY, Garcia CV. Evaluation of the protein quality of Porphyridium cruentum. J Appl Phycol 2013;25(2):497-501. [Google Scholar] | [Crossref]
- Ferreira AS, Mendonça I, Póvoa I, Carvalho H, Correia A, Vilanova M, Silva TH, Coimbra MA, Nunes C. Impact of growth medium salinity on galactoxylan exopolysaccharides of Porphyridium purpureum. Algal Res 2021:59. [Google Scholar] | [Crossref]
- Cepoi L, Rudi L, Chiriac T, Valuta A, Zinicovscaia I, Miscu V, Rudic V. Silver nanoparticles as stimulators in biotechnology of Porphyridium cruentum. Cham: Springer International Publishing; 2022. [Google Scholar]
- Sari YW, Sanders JPM, Bruins ME. Techno-economical evaluation of protein extraction for microalgae biorefinery. IOP Conf Ser Earth Environ Sci 2016;31:012034. [Google Scholar] | [Crossref]
- Wang J, Wu Y, Zhou T, Feng Y, Li L. Common factors and nutrients affecting intestinal villus height:A review. Anim Biosci 2025;38(8):1557-69. [Google Scholar] | [Crossref]
- Gatlin DM, Barrows FT, Brown P, Dabrowski K, Gaylord TG, Hardy RW. Expanding the utilization of sustainable plant products in aquafeeds:a review. Aquac Res 2007;38:551-79. [Google Scholar] | [Crossref]
- Martınez-Jeronimo F, Rodrıguez-Estrada J, Villasenor-Cordova R. Effect of culture density and volume on Moina micrura (Kurz, 1874) reproduction, and sex ratio in the progeny. Hydrobiologia 2027;594:69-73. [Google Scholar] | [Crossref]
- Kontara EK, Suryaningrum LH, Saputra A, Murniasih S, Emmawati L, Taukhid T. Review of dietary strategies for glass eels (Anguilla sp.):impact of plant-based protein diets on growth, immunity, and intestinal development. Aquac Aquarium Conserv Legis. 2025;18(2):566-86. [Google Scholar] | [Crossref]
- Ruiz de Ybáñez MR, Del Río L, Flores-Flores C, Muñoz P, Berriatua E, Rubio S, Martínez-Carrasco C. Monitoring for Anguillicoloides crassus, Anguillid herpesvirus 1, aquabirnavirus EVE and rhabdovirus EVEX in the European eel population of southern Spain. J Fish Dis 2023;46(4):417-31. [Google Scholar] | [Crossref]
- Borowitzka MA. High-value products from microalgae—their development and commercialisation. J Appl Phycol 2013;25:743-56. [Google Scholar] | [Crossref]
- Spolaore P, Cassan CJ, Duran E, Isambert A. Commercial applications of microalgae. J Biosci Bioeng 2026;101(2):87-96. [Google Scholar] | [Crossref]
- Verschuere L, Rombaut G, Sorgeloos P, Verstraete W. Probiotic bacteria as biological control agents in aquaculture. Microbiol Mol Biol Rev 2000;64(4):655-71. [Google Scholar] | [Crossref]
- Romadhona EI, Widantara H, Aslia A, Megawati N, Rahmat A, Ardiansyah. First record of Anguillid herpesvirus 1linked to a mass mortality event in shortfin eel (Anguilla bicolor) in Indonesia. J Med Vet 2025;8(1):134-43. [Google Scholar] | [Crossref]
- van Beurden SJ, Engelsma MY, Roozenburg I, Voorbergen-Laarman MA, van Tulden PW, Kerkhoff S. Viral diseases of wild and farmed European eel, Anguilla anguilla with particular reference to the Netherlands. Dis Aquat Organ 2012;101:69-86. [Google Scholar] | [Crossref]
- Crook V, Nakamura M. Glass eels:Assessing supply chain and market impacts of a CITES listing on Anguilla species. TRAFFIC Bull 2013;25(1):24-30. [Google Scholar] | [Crossref]
- Nguyen TT, Jin Y, Kiełpinska J, Bergmann SM, Lenk M, Panicz R. Detection of Herpesvirus anguillae (AngHV-1) in European eel Anguilla anguilla (L.) originating from Northern Poland assessment of suitability of selected diagnostic methods. J Fish Dis 2017;40:1717-23. [Google Scholar] | [Crossref]
- Park SW, Jung EB, Kim DW. Outbreak of Anguillid herpesvirus-1 (AngHV-1) infection in cultured shortfin eel (Anguilla bicolor) in Korea. J Fish Pathol 2012;25:151-8. [Google Scholar] | [Crossref]
- Panicz R, Eljasik P, Nguyen TT, Thi KTV, Hoang DV. First detection of Herpesvirus anguillae (AngHV-1) associated with mortalities in farmed giant mottled eel (Anguilla marmorata) in Vietnam. J Fish Dis 2021;44(6):847-52. [Google Scholar] | [Crossref]
- Guo R, Zhang Z, He T, Li M, Zhuo Y, Yang X, Fan H, Chen X. Isolation and identification of a new isolate of Anguillid herpesvirus 1from farmed American eels (Anguilla rostrata) in China. Viruses;2022(14):2722. [Google Scholar] | [Crossref]
- Crook V, Nakamura M. Glass eels:Assessing supply chain and market impacts of a CITES listing on Anguilla species. TRAFFIC Bull 2013;25(1):24-30. [Google Scholar] | [Crossref]
- Prasetiyo DE, Taukhid, Andriyanto S, Azril A, Ahyani N, Hermawati S, Ana A. Identification of microorganisms (Parasites and Bacteria) which are potential as pathogenic agent in glass eel of Anguilla spp. at the eel capture location, Sukabumi, West Java. J Aquac Sci 2021;6:170-9. [Google Scholar] | [Crossref]