

ABSTRACT
Background and Aim: The African Houbara bustard (Chlamydotis undulata), a species of high conservation concern, is maintained through intensive captive breeding programs where bacterial diseases may compromise productivity and reintroduction success. Escherichia coli is frequently implicated in reproductive failure, early mortality, and septicemic conditions; however, integrated phenotypic and genotypic characterization of antimicrobial resistance and virulence across life stages in this species remains scarce. This study aimed to determine the prevalence of E. coli across different life stages of captive African Houbara bustards, characterize multidrug-resistant (MDR), extensively drug-resistant (XDR), and extended-spectrum β-lactamase (ESBL) phenotypes, and profile avian pathogenic E. coli (APEC)–associated virulence genes within a One Health framework.
Materials and Methods: A cross-sectional diagnostic investigation was conducted during the 2025 production season. A total of 110 samples were collected from early embryonic death (n = 50), chicks (n = 25), juveniles (n = 25), and adult females (n = 10). Isolation and identification of E. coli were performed using conventional bacteriology and the VITEK®2 system. Antimicrobial susceptibility testing was carried out using disk diffusion and VITEK®2 AST, with resistance patterns classified according to international MDR and XDR definitions. Phenotypic ESBL detection was undertaken, and a multiplex real-time polymerase chain reaction (PCR) assay targeting 15 APEC-associated virulence genes was applied to representative isolates.
Results: E. coli was isolated from 62.7% (69/110) of samples, with the highest prevalence observed in juveniles (84%) and chicks (80%). All isolates were resistant to ampicillin and fluoroquinolones, while complete susceptibility was observed to fosfomycin, aminoglycosides, and chloramphenicol. MDR was detected in 96.9% of tested isolates, and one XDR isolate resistant to 15 antibiotics was identified. ESBL-producing E. coli were detected in juveniles (14%) and early embryonic death samples (9%). Virulence profiling revealed a high gene burden, with most isolates harboring ≥10 virulence genes, particularly those associated with iron acquisition, protectins, and invasion.
Conclusion: Captive African Houbara bustards harbor highly virulent MDR and ESBL-producing E. coli across life stages, posing significant conservation, veterinary, and public health risks. Strengthened biosecurity, prudent antimicrobial stewardship, and integrated One Health surveillance are essential to improve breeding success and safeguard reintroduction programs.
Keywords: African Houbara bustard, antimicrobial resistance, avian pathogenic Escherichia coli, captive breeding, extended-spectrum beta-lactamase, multidrug resistance, One Health, virulence genes.
INTRODUCTION
Of the 3800 tropical avian species worldwide, a substantial proportion has been reported as endangered or threatened [1]. According to the International Union for Conservation of Nature (IUCN) [2], the African Houbara bustard (Chlamydotis undulata) is listed on the IUCN Red List. The decline in Houbara bustard populations is primarily attributed to overhunting and habitat loss, particularly because this species is a traditional game bird for Arab falconers. The conservation of these birds is therefore imperative, and captive breeding programs play a critical role in maintaining healthy populations, reinforcing wild stocks, and safeguarding against potential population collapse [2–5]. In 2013, a captive breeding program was established at the International Foundation for Ecological Research (IFER) in Errachidia, Morocco, with the objective of conserving and reintroducing this species into its natural habitat.
Several factors, including genital infections such as salpingitis, peritonitis, and salpingo-peritonitis, can adversely compromise the reproductive performance of females in captive breeding systems [6]. Similar conditions have been documented in poultry, with Escherichia coli being one of the most frequently isolated pathogens [7, 8]. Zoonotic and pathogenic bacteria have been reported in wild birds [9], including the Houbara bustard [10]. E. coli is a major etiological agent of omphalitis, yolk sac infection, and early mortality in young chicks [11–13]. Consequently, the emergence of antimicrobial-resistant bacteria represents one of the most significant challenges facing veterinary medicine and public health [14, 15]. The escalating threat of antibiotic resistance can substantially affect both sectors by restricting the effectiveness and availability of antimicrobial therapies [16]. Inappropriate and indiscriminate use of antibiotics is a key factor exacerbating this problem [17]. Notably, antibiotic-resistant bacteria have also been detected in wild animals inhabiting remote areas with minimal or no direct exposure to antimicrobial agents [18, 19]. The introduction of captive-bred birds into wild populations has been identified as a potential route for the dissemination of antibiotic-resistant bacteria [20, 21].
Since the 2000s, resistance among Enterobacteriaceae to third- and fourth-generation cephalosporins has increasingly been reported, largely due to the production of extended-spectrum β-lactamases, resulting in limited treatment options for affected infections [22–24]. Furthermore, the interaction between virulence factors and antibiotic resistance in E. coli influences pathogenicity, disease severity, bacterial survival, and therapeutic outcomes, thereby posing additional challenges for effective disease control [25, 26].
Despite the conservation importance of C. undulata, current knowledge on bacterial diseases affecting captive breeding populations remains fragmented and largely descriptive. Previous reports have primarily focused on clinical or pathological findings, with limited integration of phenotypic resistance profiles and genotypic virulence characteristics of E. coli. In particular, comprehensive investigations that simultaneously address multidrug-resistant (MDR), extensively drug-resistant (XDR), and extended-spectrum β-lactamase (ESBL) phenotypes, and APEC-associated virulence gene burdens across different life stages are lacking in conservation-managed bustards. Moreover, age-specific patterns of resistance and virulence, especially in early embryonic death, chicks, and juveniles, remain poorly understood, limiting inference on transmission dynamics within closed breeding systems. The absence of such integrative data represents a critical gap in conservation medicine, as it constrains evidence-based biosecurity planning, antimicrobial stewardship, and One Health risk assessment related to the release of captive-bred birds into wild populations.
The present study aimed to generate an integrated phenotypic and genotypic characterization of E. coli isolated from captive C. undulata across multiple life stages. Specifically, the objectives were to determine the prevalence of E. coli in early embryonic death, chicks, juveniles, and adult females; characterize MDR, XDR, and ESBL phenotypes; assess APEC-associated virulence gene profiles using multiplex real-time polymerase chain reaction (PCR); and explore potential transmission patterns within the captive breeding system. By addressing these objectives, the study seeks to provide baseline evidence to support targeted disease control, rational antimicrobial use, and One Health–oriented conservation strategies for endangered bustard populations.
MATERIALS AND METHODS
Ethical approval
All samples were obtained from routine diagnostic postmortem examinations conducted at the IFER, Errachidia, Morocco. No live birds were handled or experimentally manipulated. Adult females, chicks, and juveniles died naturally or were submitted for necropsy as part of standard veterinary investigations, while egg samples originated from routine incubation monitoring. As no additional interventions were performed, formal ethical approval was not required. Institutional management provided a written exemption, and all procedures complied with national regulations and institutional animal welfare guidelines.
Study period and location
This study was conducted during the 2025 production season (encompassing the breeding and rearing cycle from approximately January to June, aligned with the species’ natural reproductive period under captive management) at the IFER captive breeding facility in Errachidia, Morocco. The facility is located in a semi-arid region (coordinates approximately 31.93°N, 4.42°W) and serves as a dedicated center for the conservation and reintroduction of the African Houbara bustard. All sampling, necropsies, and laboratory analyses were performed on-site or at affiliated laboratories within the IFER premises to ensure biosecurity and compliance with conservation protocols.
Study design and sampling strategy
This study was designed as a cross-sectional diagnostic investigation conducted during the 2025 production season. A purposive sampling strategy was used, in which all cases meeting the inclusion criteria were included in the study. In total, 110 samples were collected and categorized into distinct stages of the reproductive and early life cycle of C. undulata: early embryonic death (n = 50), chicks (n = 25), juveniles (n = 25), and adult females (n = 10). The selected sample size reflected the range of age categories encountered and corresponded to routine diagnostic submissions received during the study period to support bacteriological and molecular analyses.
Clinical examination and necropsy procedures
A total of 110 cases were examined, comprising 10 adult females, 25 chicks, 25 juveniles, and 50 early embryonic death samples. Among adult females, six cases of sudden death were recorded, while four additional birds exhibited lethargy and anorexia prior to death, including two cases complicated by dystocia and oviductal impaction. Of the chicks examined, 15 died suddenly, and 10 displayed clinical signs of anorexia and lethargy before death. In the juvenile group, 12 birds died suddenly without overt clinical signs, whereas 13 showed systemic illness characterized by ruffled feathers, marked lethargy, and anorexia. Fifty egg samples exhibiting early embryonic mortality at 5–7 days of incubation were also included.
Necropsies were performed by licensed veterinarians following established standard protocols [27]. Postmortem examination revealed macroscopic lesions consistent with E. coli septicemia, including fibrinous pericarditis, perihepatitis, airsacculitis, exudate accumulation, peritonitis, and visceral adhesions.
Sample collection and processing
All samples were collected aseptically from internal organs, including the heart, liver, lung, and spleen, as well as oviduct and ovary tissues from adult females when indicated by clinical and pathological findings. Eggshell surfaces were decontaminated by immersion in 70% ethanol for 5–10 s, air-dried, aseptically cracked, and the internal contents pooled [28]. Samples were transported to the IFER laboratory in sterile stomacher bags under cooled conditions. Tissue samples were pooled and processed for bacterial screening and isolation.
Bacteriological isolation and identification of E. coli
Isolation and identification of E. coli were performed following previously described methods [29]. Briefly, pooled organ samples were pre-enriched in peptone water and incubated aerobically at 37°C for 24 h, followed by streaking onto MacConkey agar and EMB agar (Oxoid, Hampshire, UK) and incubation at 37°C for 24–48 h. Presumptive colonies were subcultured to obtain pure isolates. Biochemical identification was carried out using the VITEK®2 system (bioMérieux SA, Marcy-l’Étoile, France). Bacterial suspensions were adjusted to 0.5 McFarland, loaded into VITEK®2 GN cards (Ref. No. 21341), and processed according to the manufacturer’s instructions to obtain species-level identification within the Enterobacteriaceae family.
Antimicrobial susceptibility testing and ESBL phenotyping
Antimicrobial susceptibility testing was performed on a representative subset of 32 isolates using VITEK®2 GN97 cards (Ref. No. 42000, bioMérieux SA, Marcy-l’Étoile, France) to assess susceptibility to 18 antimicrobial agents and screen for ESBL production. The disk diffusion method was additionally applied following EUCAST guidelines and the latest VITEK®2 clinical breakpoint tables (version 14.0, 2024) [30]. E. coli ATCC 25922 was used as the quality control strain. Resistance patterns were classified according to international definitions [31] as MDR (resistance to ≥1 agent in ≥3 antimicrobial classes) and XDR (resistance to ≥1 agent in all but one or two classes).
Genomic DNA extraction and quality assessment
Genomic DNA was extracted from 20 overnight bacterial cultures grown to an optical density of 1.0–1.2 at OD600. Cell pellets were obtained by centrifugation at 8,000 × g for 5 min and processed using the Kylt® DNA/RNA Kit (Kybio Co., Ltd., Shenzhen, China) following the manufacturer’s instructions. DNA concentration and purity were assessed using a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), with acceptable purity defined as an A260/A280 ratio of 1.8–2.0.
Multiplex real-time PCR for APEC virulence genes
Virulence-associated genes were detected using the Kylt® APEC qPCR Kit (Kybio Co., Ltd., Shenzhen, China), targeting 15 APEC-related genes, including adhesins (papC, tsh), invasion (ibeA), iron acquisition (iucD, iutA, irp2, iroN), toxins (astA, vat, F11, hlyF), protectins (sitA, cvi/cva, iss), and outer membrane protease (ompT). Four multiplex reactions (APEC1–APEC4) were prepared in a final volume of 20 µL containing 16 µL master mix and 4 µL template DNA. Amplification was performed using a QuantStudio™ 5 Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA) with cycling conditions of 95°C for 10 min, followed by 42 cycles of 95°C for 15 s and 60°C for 60 s. Samples with Ct ≤ 30 were considered positive. Negative controls were included in all runs. Targeted genes are summarized in Table 1.
Table 1. Targeted virulence genes in four multiplex polymerase chain reaction (PCR) mixes for avian pathogenic Escherichia coli.
| Real-time PCR mix | HEX (target gene) | FAM (target gene) | Cy5 (target gene) | TXR (target gene) |
|---|---|---|---|---|
| APEC1 | Internal Control | papC gene | tsh gene | irp2 gene |
| APEC2 | iss gene | iucD gene | F11 gene | astA gene |
| APEC3 | ibeA gene | vat gene | cvi/cva gene | iutA gene |
| APEC4 | iroN gene | hlyF gene | ompT gene | sitA gene |
Statistical analysis
Data were analyzed using Microsoft Excel® and SPSS® version 26 (IBM Corp., Armonk, NY, USA). Descriptive statistics were used to calculate frequencies and percentages of E. coli isolates and virulence genes. Fisher’s exact test was applied to compare prevalence among groups due to small sample sizes. Statistical significance was set at p ≤ 0.05. Confidence intervals are presented in Table 2.
Table 2. Prevalence of Escherichia coli isolates from different life stages of Houbara bustard.
| Sample sources | Organs/tissue | Number of samples collected | No. of positive samples | % Positive (95% CI) | Interpretation |
|---|---|---|---|---|---|
| Dead females | Heart-liver-lung-spleen-oviduct | 10 | 6 | 60 (29.6–90.4) | Small sample size is less precise |
| Egg (EED) | Egg content | 50 | 22 | 44 (30.3–57.7) | Larger sample makes it more reliable |
| Chicks | Heart-liver-lung-spleen | 25 | 20 | 80 (64.3–95.7) | Strong and clear results |
| Juvenile | Heart-liver-lung-spleen | 25 | 21 | 84 (69.6–98.4) | Strong and clear results |
| Total | — | 110 | 69 | 62.7 (53.7–71.7) | Large sample and precise result |
EED = Early embryonic death, CI = Confidence interval.
RESULTS
Isolation and identification of E. coli
E. coli was isolated from 69 of 110 cases (62.7%). The highest isolation rate was observed in juveniles (84%), followed by chicks (80%), adult females (60%), and early embryonic death (44%). Sixty-two clinical isolates were identified as E. coli using the VITEK®2 system, with identification confidence ranging from 96% to 99%. Minor deviations in expected biochemical reactions (PHOS-81 and dTAG-22) were detected in a limited number of isolates; however, overall identification confidence remained high. Confidence intervals were calculated for each group.The prevalence of E. coli isolates across different life stages of C. undulata is summarized in Table 2.
Phenotypic clustering and antimicrobial resistance profiles
Two clustering patterns were identified: phenotypic clustering based on antimicrobial susceptibility profiles and genotypic clustering based on virulence gene distribution. Figure 1 presents a heat map illustrating resistance percentages among E. coli isolates recovered from early embryonic death, chicks, juveniles, and adult females.
Heatmap showing the percentage resistance of Escherichia coli isolates to individual antibiotics, stratified by sample source (early embryonic death, chicks, juveniles, and adult females) in captive African Houbara bustards.
Isolates originated from eggs (n = 8), chicks (n = 7), juveniles (n = 12), and adult females (n = 5). No resistance was detected to fosfomycin, amikacin, gentamicin, neomycin, or chloramphenicol. In contrast, all isolates exhibited resistance to enrofloxacin (ENR) and ampicillin (AMP).
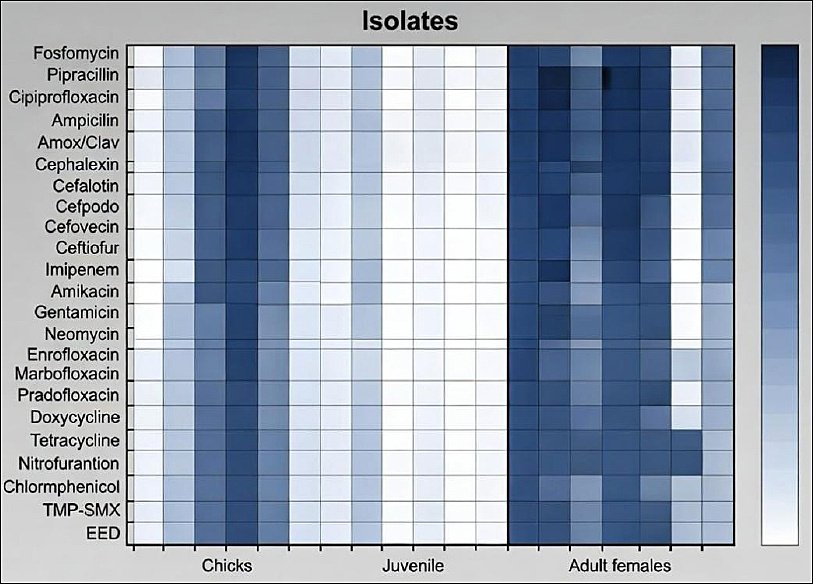
Figure 1. Heatmap showing the percentage resistance of Escherichia coli isolates to individual antibiotics, stratified by sample source (early embryonic death, chicks, juveniles, and adult females) in captive African Houbara bustards.
Distribution of multidrug-resistant phenotypes
The distribution of phenotypic antibiotic resistance patterns is shown in Table 3. A total of 25 distinct multidrug-resistant (MDR) patterns were identified, ranging from resistance to 3 to 15 antibiotics. The most frequent resistance profile consisted of AMP, marbofloxacin (MAR), pradofloxacin (PRA), doxycycline (DOX), tetracycline (TET), nitrofurantoin (NIT), and trimethoprim–sulfamethoxazole (TMP-SMX) and was observed in three isolates. Several additional resistance profiles involved combinations of 8–10 antibiotics. One isolate demonstrated resistance to 15 antibiotics, indicating an extensively drug-resistant (XDR) phenotype. AMP, fluoroquinolones, TET, NIT, and TMP-SMX were the most frequently represented antimicrobial classes across resistance patterns. The distribution of MDR among sample groups is presented in Figure 2. Of the 32 E. coli isolates examined, 31 (96.9%) were classified as MDR. All isolates from chicks, juveniles, and adult females exhibited 100% MDR. Only one isolate from the early embryonic death group was non-MDR, resulting in an MDR prevalence of 87.5% in that group.
Distribution of multidrug-resistant (MDR) Escherichia coli isolates across sample groups (early embryonic death, chicks, juveniles, and adult females).
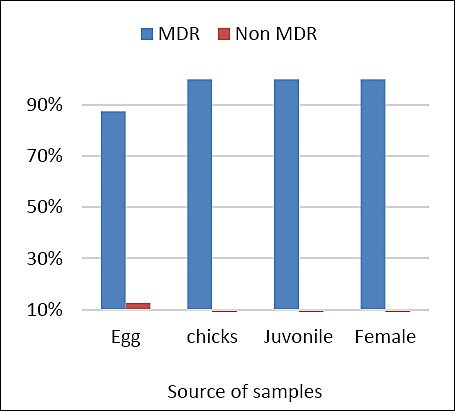
Figure 2. Distribution of multidrug-resistant (MDR) Escherichia coli isolates across sample groups (early embryonic death, chicks, juveniles, and adult females).
Table 3. Distribution of phenotypic patterns of antibiotic resistance in Escherichia coli among different Houbara bustard samples.
| Pattern no. | Resistant pattern (antibiotics) | Number of isolates with pattern | Number of antibiotics in pattern |
|---|---|---|---|
| 1 | CIP + AMP + AMC | 1 | 3 |
| 2 | AMP + MAR + DOX + TET + NIT + TMP-SMX | 1 | 6 |
| 3 | AMP + NEO + MAR + DOX + NIT + TMP-SMX | 1 | 6 |
| 4 | CIP + AMP + MAR + DOX + TET + NIT | 1 | 6 |
| 5 | AMP + CFV + MAR + PRA + TET + NIT | 1 | 6 |
| 6 | AMP + MAR + PRA + DOX + TET + NIT + TMP-SMX | 3 | 7 |
| 7 | CIP + AMP + MAR + PRA + TET + NIT + TMP-SMX | 1 | 7 |
| 8 | AMP + AMC + MAR + PRA + DOX + TET + NIT + TMP-SMX | 2 | 8 |
| 9 | CIP + AMP + AMC + MAR + DOX + TET + NIT + TMP-SMX | 2 | 8 |
| 10 | CIP + AMP + AMC + MAR + PRA + DOX + TET + NIT | 2 | 8 |
| 11 | CIP + AMP + MAR + PRA + DOX + TET + NIT + TMP-SMX | 1 | 8 |
| 12 | CIP + AMP + AMC + CFV + MAR + PRA + DOX + TET + NIT | 1 | 9 |
| 13 | CIP + AMP + AMC + CFX + MAR + DOX + TET + NIT + TMP-SMX | 1 | 9 |
| 14 | CIP + AMP + AMC + CFX + MAR + PRA + DOX + TET + NIT | 1 | 9 |
| 15 | CIP + AMP + AMC + CPD + MAR + PRA + DOX + NIT + TMP-SMX | 1 | 9 |
| 16 | CIP + AMP + AMC + MAR + PRA + DOX + TET + NIT + TMP-SMX | 1 | 9 |
| 17 | PRL + AMP + AMC + MAR + PRA + DOX + TET + NIT + TMP-SMX | 1 | 9 |
| 18 | PRL + AMP + CPD + MAR + PRA + DOX + TET + NIT + TMP-SMX | 1 | 9 |
| 19 | PRL + CIP + AMP + AMC + MAR + PRA + DOX + TET + NIT | 1 | 9 |
| 20 | CIP + AMP + AMC + CFX + CPD + MAR + DOX + TET + NIT + TMP-SMX | 2 | 10 |
| 21 | CIP + AMP + AMC + CFX + CPD + MAR + PRA + DOX + TET + NIT | 1 | 10 |
| 22 | PRL + CIP + AMP + AMC + MAR + PRA + DOX + TET + NIT + TMP-SMX | 2 | 10 |
| 23 | AMP + AMC + CFX + CPD + CFV + MAR + PRA + DOX + TET + NIT + TMP-SMX | 1 | 11 |
| 24 | PRL + CIP + AMP + AMC + CPD + MAR + PRA + DOX + TET + NIT + TMP-SMX | 1 | 11 |
| 25 | CIP + AMP + AMC + CFX + CLT + CPD + CFV + CEF + IPM + MAR + PRA + DOX + TET + NIT + TMP-SMX | 1 | 15 |
AMP = Ampicillin, AMC = Amoxicillin-Clavulanic Acid, PRL = Piperacillin, CIP = Ciprofloxacin, CLT = Cefalotin, CPD = Cefpodoxime, CFV = Cefovecin, CFX = Cefalexin, CEF = Ceftiofur, IPM = Imipenem, NEO = Neomycin, MAR = Marbofloxacin, PRA = Pradofloxacin, DOX = Doxycycline, TET = Tetracycline, NIT = Nitrofurantoin, TMP-SMX = Trimethoprim-Sulfamethoxazole.
Life stage–specific MDR patterns
Figure 3 illustrates the distribution and frequency of MDR patterns across life stages. The most predominant resistance combination consisted of β-lactams, fluoroquinolones, TET, NIT, and sulfonamides in chicks (71.5%) and eggs (75%). Adult females exhibited a higher proportion (60%) of resistance to β-lactams, fluoroquinolones, TET, and NIT. Juvenile isolates showed multiple resistance combinations. These findings indicate substantial variation in MDR patterns among life stages, although overall MDR prevalence remained consistently high (96.9%) across all groups.
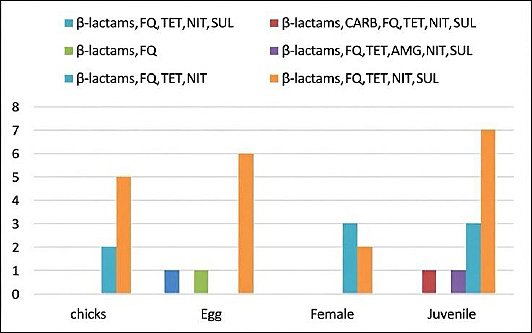
Figure 3. Distribution of multidrug-resistant patterns across life stages of Houbara bustards. β-lactams = Beta-lactams, FQ = Fluoroquinolones, TET = Tetracyclines, SUL = Sulfonamides, NIT = Nitrofurans, AMG = Aminoglycosides, CARB = Carbapenems.
Statistical analysis of antimicrobial resistance
Statistical analysis of resistance rates by antimicrobial class is presented in Table 4. Due to small sample sizes in certain groups, Fisher’s exact test was applied instead of the chi-square test. No statistically significant differences in resistance rates were detected among early embryonic death, chicks, juveniles, and adult females (p > 0.05), largely due to uniform resistance and susceptibility patterns across most antimicrobial classes.
Table 4. Statistical analysis of antibiotic resistance by families across Houbara bustard groups using Fisher’s exact test and Chi-square test.
| Antibiotic family | EED (n=8) | Chicks (n=7) | Juveniles (n=12) | Adult females (n=5) | Test used | p-value | Significance |
|---|---|---|---|---|---|---|---|
| Aminoglycosides | 0% | 0% | 0% | 0% | N/A (all sensitive) | N/A | NS |
| Fluoroquinolones | 100% | 100% | 100% | 100% | N/A (all resistant) | N/A | NS |
| Beta-lactams | 100% | 100% | 100% | 100% | N/A (all resistant) | N/A | NS |
| Carbapenem | 0% | 0% | 0% | 0% | N/A (all sensitive) | N/A | NS |
| Phosphonic acid | 0% | 0% | 0% | 0% | N/A (all sensitive) | N/A | NS |
| Tetracyclines | 87.5% | 100% | 100% | 100% | Fisher’s exact test | 0.33 | NS |
| Nitrofurans | 87.5% | 100% | 100% | 100% | Fisher’s exact test | 0.33 | NS |
| Phenicol | 0% | 0% | 0% | 0% | N/A (all sensitive) | N/A | NS |
| Folate pathway inhibitors | 87.5% | 71.4% | 75% | 40% | Fisher’s exact test | 0.20 | NS |
EED = Early embryonic death, NS = Non-significant, N/A = Non-applicable.
Prevalence of ESBL-producing E. coli
The prevalence of extended-spectrum β-lactamase–producing E. coli across life stages is shown in Figure 4. ESBL-producing isolates were detected in 14% of juvenile samples and 9% of egg samples, indicating stage–specific occurrence and highlighting the need for targeted monitoring during early developmental stages.
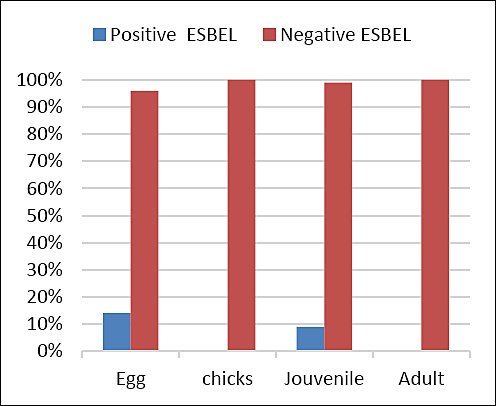
Figure 4. Prevalence of extended-spectrum β-lactamase-producing (ESBL) Escherichia coli in Houbara bustards across different life stages (early embryonic death, chicks, juveniles, and adult females).
Virulence gene profiles of APEC isolates
The prevalence of avian pathogenic E. coli–associated virulence genes is summarized in Table 5. Most isolates harbored irp2, iss, iucD, iutA, ompT, and sitA, which were consistently detected across isolates from chicks, early embryonic death, juveniles, and adult females. In contrast, papC and tsh were absent in all isolates. Greater virulence gene diversity was observed in chick and egg isolates compared with juvenile isolates.
Gene prevalence and patterns of gene absence by isolate group (n = 20) are illustrated in Figure 5. A conserved core gene set, including protectin- and invasion-associated genes, was detected in 100% (20/20) of isolates, indicating their essential role in bacterial survival and pathogenicity. Iron acquisition genes exhibited the highest collective prevalence (17/20–20/20), emphasizing their importance in nutrient acquisition. Conversely, adhesin and toxin genes were detected at low frequencies (0/20–3/20), reflecting heterogeneity in pathogenic potential.
Gene prevalence and absence patterns for 15 avian pathogenic Escherichia coli-associated virulence genes among 20 selected E. coli isolates, grouped by source (n = 20). Prevalence numbers (x/20) = Total number of positive isolates (isolates possessing the gene). The presence of a group letter next to a gene indicates absence of that gene in the corresponding group (CH = chicks, EED = early embryonic death, F = adult female, and J = juvenile).
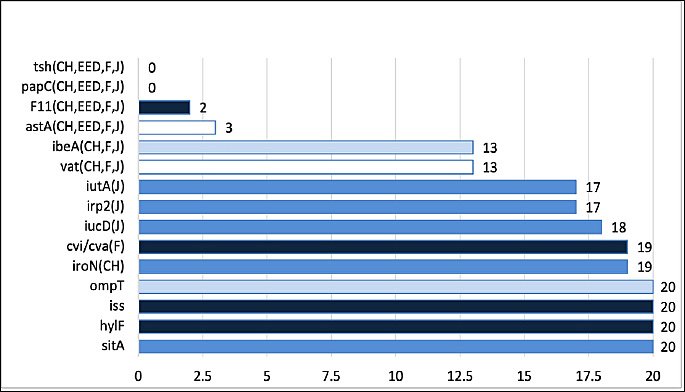
Figure 5. Gene prevalence and absence patterns for 15 avian pathogenic Escherichia coli-associated virulence genes among 20 selected E. coli isolates, grouped by source (n = 20). Prevalence numbers (x/20) = Total number of positive isolates (isolates possessing the gene). The presence of a group letter next to a gene indicates absence of that gene in the corresponding group (CH = chicks, EED = early embryonic death, F = adult female, and J = juvenile).
Table 5. Prevalence of APEC-associated 15 virulence genes in selected Escherichia coli isolates recovered from early embryonic death, chicks, juveniles, and adult female African Houbara bustards.
| Isolate ID | Adhesins | Iron Acquisition | Toxins | Protectins | Invasins | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
| |||||||||||
| papC | tsh | irp2 | iucD | iutA | sitA | iroN | astA | vat | F11 | hlyF | iss | cvi/cva | ompT | ibeA | |
| Chick-1 | – | – | + | + | + | + | + | – | + | – | + | + | + | + | + |
| Chick-2 | – | – | + | + | + | + | + | – | + | – | + | + | + | + | + |
| Chick-3 | – | – | + | + | + | + | + | – | – | + | + | + | + | + | – |
| Chick-4 | – | – | + | + | + | + | + | – | + | – | + | + | + | + | + |
| Chick-5 | – | – | + | + | + | + | – | – | – | – | + | + | + | + | – |
| EED-1 | – | – | + | + | + | + | + | – | + | – | + | + | + | + | + |
| EED-2 | – | – | + | + | + | + | + | + | + | – | + | + | + | + | + |
| EED-3 | – | – | + | + | + | + | + | – | + | – | + | + | + | + | + |
| EED-4 | – | – | + | + | + | + | + | + | + | – | + | + | + | + | + |
| EED-5 | – | – | + | + | + | + | + | – | + | – | + | + | + | + | + |
| Female-1 | – | – | + | + | + | + | + | – | – | + | + | + | – | + | – |
| Female-2 | – | – | + | + | + | + | + | – | – | – | + | + | + | + | – |
| Female-3 | – | – | + | + | + | + | + | – | + | – | + | + | + | + | + |
| Female-4 | – | – | + | + | + | + | + | + | + | – | + | + | + | + | + |
| Female-5 | – | – | + | + | + | + | + | – | + | – | + | + | + | + | + |
| Juvenile-1 | – | – | – | – | – | + | + | – | – | – | + | + | + | + | – |
| Juvenile-2 | – | – | – | + | + | + | + | – | – | – | + | + | + | + | – |
| Juvenile-3 | – | – | + | + | – | + | + | – | + | – | + | + | + | + | + |
| Juvenile-4 | – | – | + | – | – | + | + | – | + | – | + | + | + | + | + |
| Juvenile-5 | – | – | – | + | + | + | + | – | – | – | + | + | + | + | – |
EED = Early embryonic death APEC = Avian pathogenic Escherichia coli (+) = Presence of the gene (–) = Absence of the gene.
Mean virulence gene burden across life stages
The mean virulence gene burden across life stages of C. undulata is shown in Figure 6. Juvenile isolates exhibited the lowest standard deviation, indicating consistent gene burden values within this group, whereas chick isolates showed the highest standard deviation, reflecting greater variability in virulence gene distribution.
Mean number of virulence genes (gene burden) carried by Escherichia coli isolates across different life stages of Houbara bustards. EED = Early embryonic death.
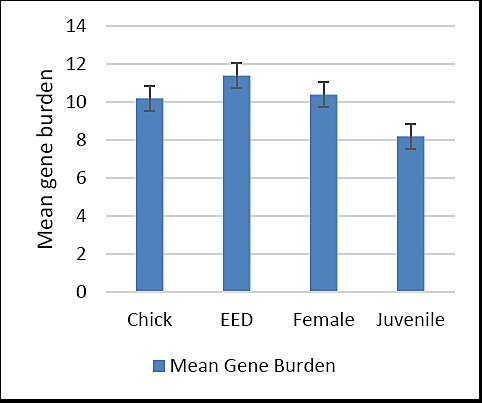
Figure 6. Mean number of virulence genes (gene burden) carried by Escherichia coli isolates across different life stages of Houbara bustards. EED = Early embryonic death.
DISCUSSION
Current knowledge gaps and study contribution
Only limited literature has addressed MDR E. coli and APEC-associated virulence factors in C. undulata, resulting in a substantial gap in understanding the pathogenic potential and epidemiology of E. coli in endangered avian species [8, 32, 33]. Consequently, the present findings represent one of the first comprehensive investigations integrating phenotypic MDR profiling with genotypic virulence characterization, including the detection of XDR isolates circulating within captive flocks [34, 35]. The combined phenotypic–genotypic framework applied in this study enabled detailed assessment of pathogen dynamics across life stages, supported by profiling of 15 virulence-associated genes [36, 37]. Isolates recovered from breeders, early embryonic death, chicks, and juveniles were associated with elevated mortality in captive African Houbara bustards (C. undulata).
Prevalence of E. coli across life stages
The overall isolation rate of E. coli (62.7%) in C. undulata was notably high. This contrasts with a previous report describing 100% prevalence in postmortem samples from Chlamydotis macqueenii [38]. The highest isolation rates were recorded in juveniles (84%) and chicks (80%), supporting the association between immature immune function and age-related susceptibility in avian species. Environmental stressors have been widely reported as predisposing factors for colibacillosis in young birds [39–41]. Isolation rates in adult females (60%) during the breeding season were consistent with previous findings identifying E. coli as the predominant pathogen in clinical reproductive disorders of captive Houbara bustards [6]. Detection of E. coli in early embryonic death samples (44%) supports the hypothesis of horizontal or vertical transmission in avian systems [42–45], although recent studies indicate that true vertical transmission in poultry remains limited, with environmental contamination playing a more dominant role [46, 47].
Antimicrobial resistance patterns and XDR emergence
Marked phenotypic resistance was observed against AMP, ENR, MAR, TET, NIT, DOX, and PRA across all isolates, in agreement with previous reports from Houbara bustards and poultry production systems [38, 42, 48, 49]. Earlier studies documented low ENR resistance in Houbara bustards and other wild birds [38, 50]; therefore, the widespread ENR resistance observed here indicates a concerning shift in resistance dynamics. Notably, PRA resistance was identified for the first time in Houbara bustards, highlighting the emergence of novel resistant phenotypes.
In contrast, complete susceptibility was observed to fosfomycin, amikacin, gentamicin, and chloramphenicol, supporting their potential role as alternative therapeutic options. These findings align with previous observations in poultry, where universal susceptibility to fosfomycin was reported [51]. Low resistance levels against cefalotin, cefovecin, ceftiofur, and neomycin in juvenile isolates further suggest limited but promising treatment alternatives for APEC infections in captive bustards.
Alarmingly, one isolate exhibited resistance to 15 antibiotics, including imipenem, indicating the emergence of XDR strains (Table 3). Similar findings have been reported in poultry populations [35]. MDR patterns were consistently distributed across all life stages, predominantly involving β-lactams, fluoroquinolones, TET, NIT, and sulfonamides (40%–75%), corroborating earlier reports linking resistance gene accumulation with treatment failure risks [32, 33].
ESBL-producing E. coli and public health implications
The indiscriminate use of antibacterial agents across human, veterinary, and agricultural sectors has been implicated in the global dissemination of ESBL-producing bacteria [52, 53]. Sub-therapeutic antibiotic exposure in poultry has been proposed as a driver for environmental dissemination of ESBL-producing E. coli, posing substantial public health risks [22, 54]. While transmission of ESBL-producing E. coli between poultry products and humans has been documented [55], data regarding transmission from captive Houbara bustards to personnel remain unavailable.
In the present study, ESBL-producing E. coli were detected primarily during early life stages, with positivity rates of 15% in early embryonic death and 10% in juveniles. These findings are consistent with previous observations suggesting early horizontal exposure or breeder-associated transmission [36]. Juveniles, due to immunological immaturity, may facilitate persistence and dissemination of resistant strains [32, 33]. The absence of statistically significant differences among age groups (p > 0.05) supports flock-level dissemination of MDR E. coli, reinforcing the need for stringent biosecurity during incubation and early rearing.
Virulence gene distribution and life stage specificity
APEC isolates are characterized by diverse virulence determinants, including adhesins, toxins, iron acquisition systems, and invasion-associated factors [56, 57]. Distinct life stage–specific virulence patterns were identified in captive C. undulata. Early embryonic death and chick isolates exhibited the highest virulence gene burdens, with most isolates harboring ≥10 genes, including iss, iucD, iutA, ompT, hlyF, and sitA. Similar gene burden–based pathogenicity classifications have been reported in poultry [58], although others have questioned the discriminatory power of virulence gene profiling alone [59].
Juvenile isolates displayed intermediate virulence profiles, potentially reflecting a transitional phase with reduced pathogenic potential. Adult female isolates retained core virulence genes, such as iss, iutA, and ompT, but exhibited lower overall gene richness, consistent with previous reports in Houbara bustards [6]. Reduced virulence gene prevalence may be associated with enhanced immune competence or cumulative environmental exposure.
The absence of certain adhesion-associated genes further emphasizes that E. coli pathogenicity is multifactorial and influenced not only by virulence gene content but also by host immunity, susceptibility, and environmental and management conditions [8, 33, 60, 61].
CONCLUSION
This study demonstrated a high burden of E. coli infection (62.7%) across life stages of captive C. undulata, with the highest prevalence observed in juveniles and chicks. An alarmingly high proportion of isolates exhibited MDR (96.9%), with the detection of an XDR isolate confirming the emergence of highly resistant strains within the breeding system. ESBL-producing E. coli were predominantly detected during early life stages, supporting early acquisition and flock-level dissemination. Genotypic analysis revealed that most isolates carried multiple APEC-associated virulence genes, with early embryonic death and chick isolates harboring the highest gene burdens, particularly iss, iucD, iutA, ompT, hlyF, and sitA.
The coexistence of MDR, XDR, ESBL phenotypes, and high virulence gene loads in E. coli poses significant challenges for disease control, treatment efficacy, and conservation success in captive C. undulata. These findings highlight the urgent need for strengthened biosecurity, rational antimicrobial stewardship, and routine resistance surveillance within captive breeding facilities. Targeted hygiene interventions during incubation and early rearing are particularly critical to reduce early-life exposure. From a One Health perspective, the presence of highly resistant and virulent strains underscores potential risks to personnel, surrounding wildlife, and the environment.
Key strengths include the integrated phenotypic–genotypic approach, simultaneous assessment of MDR, XDR, ESBL, and APEC profiles, and life stage–specific analysis across the production cycle. The application of a 15-gene virulence panel enabled high-resolution characterization of pathogenic potential, providing one of the most comprehensive datasets currently available for E. coli in conservation-managed bustards.
The limited number of adult female samples may restrict extrapolation of findings for this group. In addition, reliance on targeted virulence profiling without whole-genome sequencing constrained deeper resolution of resistance mechanisms, plasmid structures, and transmission pathways. The focus on diagnostic cases may also overrepresent clinically severe infections.
Future studies should incorporate whole-genome sequencing to resolve resistance determinants, mobile genetic elements, and transmission dynamics within and beyond breeding facilities. Longitudinal monitoring, environmental sampling, and assessment of breeder-associated transmission routes are warranted. Evaluation of non-antibiotic interventions, including vaccination strategies and probiotic-based approaches, may offer sustainable alternatives for disease mitigation.
Overall, this study provides critical evidence that captive C. undulata populations can serve as reservoirs of MDR, XDR, ESBL-producing, and highly virulent E. coli. Integrating rigorous biosecurity, antimicrobial stewardship, and One Health–aligned surveillance is essential to safeguard conservation outcomes, animal health, and public health interfaces in captive breeding and reintroduction programs.
DATA AVAILABILITY
All the generated data are included in the manuscript.
AUTHOR’S CONTRIBUTIONS
The author solely contributed to the study’s conception and design, data collection, analysis and interpretation, manuscript writing, and final approval of the submitted version.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The author expresses her sincere gratitude to Mr. Khamis Morshid El-Marikhy, Director of the IFER, for his continuous support, expert guidance, and unwavering encouragement throughout this study. His deep commitment to wildlife conservation and the welfare of the African Houbara bustard was instrumental in enabling the successful completion of this research. The author is particularly thankful for his provision of essential resources, access to facilities, and strong belief in the value of this work.
The author also extends sincere thanks to the technical staff members Abil Kavukkalathil and Muhammed Arshad V. for their dedicated assistance with laboratory procedures and data management.
Special appreciation is due to Dr. Adel Abd Errahman for conducting the postmortem examinations, as well as to Biman Das Pukaysta, Dr. Palash Das, and Mr. Edris Mogany for performing the egg break tests and documenting case histories. Their skilled contributions significantly supported the progress and quality of this study. This research was funded by the International Foundation for Ecological Research (IFER), Morocco.
REFERENCES
- Díaz S, Fargione J, Chapin FI, Tilman D. Biodiversity loss threatens human well-being. PLoS Biol 2009;4:e277. [Google Scholar] | [Crossref]
- The IUCN Red List of Threatened Species:Chlamydotis undulata. International Union for Conservation of Nature 2014. [Google Scholar] | [Crossref]
- Ralls K, Meadows L. Captive breeding programs and the conservation of endangered species. Conserv Biol 2001;15:1080-1090. [Google Scholar] | [Crossref]
- Conde DA, Colchero F, Guillén J. Integrating life history and demography in conservation programs. Biol Conserv 2011;144:2562-2570. [Google Scholar] | [Crossref]
- Taylor G, Lacy R, Feistner A. Captive breeding in conservation. Zoo Biol 2017;36:127-136. [Google Scholar] | [Crossref]
- Crispo E, Smith J, Thompson R. Genital infections in captive birds:salpingitis and peritonitis. Avian Pathol 2025;54:12-25. [Google Scholar] | [Crossref]
- Landman W, Feberwee A, Mevius D. Avian colibacillosis:pathogenesis and control strategies. Avian Pathol 2013;42:403-410. [Google Scholar] | [Crossref]
- Nolan L, Barnes H, Vaillancourt J. Colibacillosis in poultry:new insights. Poult Sci 2020;99:16-29. [Google Scholar] | [Crossref]
- Dobbin G, Paul N, Gibb ZZ. Zoonotic bacteria in wild birds. J Wildl Dis 2005;41:725-733. [Google Scholar] | [Crossref]
- Stievenart C, Mohammed H. Health status of Houbara bustards in captivity. Int J Avian Sci 2004;146:203-211. [Google Scholar] | [Crossref]
- Effendi, Ramandinianto SC, Wibowo S, Fauziah I, Kusala MKJ, Fauzia KA, Furqoni AH, Raissa R. Omphalitis and yolk sac infection in poultry:etiology, pathology, and epidemiology. Vet World 2024;17(12):1050-1060. [Google Scholar] | [Crossref]
- Hermawan FA, Nadania Zega DIS, Triatjaya Y, Khairani S, Pratiwi U. Anatomical pathology features in day-old chicks with omphalitis. ARSHI Vet Lett 2024;8(3):53-54. [Google Scholar] | [Crossref]
- Shahjada F, Rahman M, Islam K. Pathogenic E. coliin poultry production. J Vet Sci 2017;18:123-130. [Google Scholar] | [Crossref]
- World Organisation for Animal Health:antimicrobial resistance report. OIE Publishing 2019. [Google Scholar] | [Crossref]
- Naghavi M, Vollset SE, Ikuta KS, Swetschinski LR. Global burden of bacterial antimicrobial resistance 1990–2021:a systematic analysis with forecasts to 2050. Lancet 2024;404(10459):1199-1226. [Google Scholar] | [Crossref]
- Global Antibiotic Resistance Surveillance Report 2025. Geneva: World Health Organization; 2025. [Google Scholar]
- Pulingam T, Rahman A, Khan S. Inappropriate antibiotic use in veterinary medicine. Antibiotics 2022;11:678. [Google Scholar] | [Crossref]
- Bartoloni A, Cutts F, Leoni S. Antibiotic resistance in remote areas. Trop Med Int Health 2004;9:467-471. [Google Scholar] | [Crossref]
- Gilliver R, Bennett M, Begon M. Antibiotic-resistant bacteria in wild animals. Environ Microbiol 1999;1:361-367. [Google Scholar] | [Crossref]
- Woodford N, Kock R. Transmission of antibiotic-resistant bacteria. J Antimicrob Chemother 1991;27:15-25. [Google Scholar] | [Crossref]
- Woodford N. Extended-spectrum beta-lactamases in Enterobacteriaceae. Clin Microbiol Infect 2000;6:460-466. [Google Scholar] | [Crossref]
- Poirel L, Madec J, Lupo A. Resistance to cephalosporins via ESBLs in Enterobacteriaceae. Front Microbiol 2018;9:2082. [Google Scholar] | [Crossref]
- Giufrè M, Accogli M, Cerquetti M. ESBL-producing Ecoliin animals and humans. Antibiotics 2021;10:125. [Google Scholar] | [Crossref]
- Cardozo M, Furlan J, Souza R. Extended-spectrum beta-lactamase producing E. coli:epidemiology and public health implications. J Glob Antimicrob Resist 2022;28:62-70. [Google Scholar] | [Crossref]
- Pan Y, Zhao F, Li X. Interplay between virulence factors and antibiotic resistance in Ecoli. Front Microbiol 2020;11:1351. [Google Scholar] | [Crossref]
- Sora V. Extraintestinal pathogenic Ecoli:virulence gene profiles and antimicrobial resistance patterns. Microorganisms 2021;9:1234. [Google Scholar] | [Crossref]
- MajóMasferrer N, Dolz Pascual R. Atlas of Avian Necropsy:Macroscopic Diagnosis Sampling (Updated Edition) 2019. [Google Scholar] | [Crossref]
- El Ftouhy FZ, Nassik S, Nacer S, Kadiri A, Charrat N, Attrassi K, Hmyene A. Bacteriological quality of table eggs in Moroccan formal and informal sector. Int J Food Sci 2022;2022:6223404. [Google Scholar] | [Crossref]
- Nolan L, Barnes H, Vaillancourt J, Abdul-Aziz T, Logue CM, Saif YM, Fadly AM, Glisson JR, McDougald LR, Nolan LK, Swayne DE. Colibacillosis. Ames, IA: Wiley-Blackwell; 2013. p. 751-805. [Google Scholar]
- Antimicrobial Resistance:Global Report on Surveillance. Geneva: World Health Organization; 2003. [Google Scholar]
- Magiorakos AP, Srinivasan A, Carey RB. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria:standard definitions for acquired resistance. Clin Microbiol Infect 2012;18:268-281. [Google Scholar] | [Crossref]
- Ewers C, Janssen T, Kiessling S, Philipp H, Wieler L. Avian pathogenic Escherichia coli (APEC). Vet Microbiol 2007;123((1–3)):91-118. [Google Scholar] | [Crossref]
- Dziva F, Stevens M. Colibacillosis in poultry:pathogenesis and strategies for control. Avian Pathol 2008;37(2):133-149. [Google Scholar] | [Crossref]
- Liang X, Wu J, Zhang Q. Multidrug-resistant Ecoliin poultry farms. Front Vet Sci 2023;10:101234. [Google Scholar] | [Crossref]
- Aworh M, Kwaga J, Okeke I. Emergence of extensively drug-resistant Ecoliin Nigerian poultry. Antimicrob Resist Infect Control 2021;10:145. [Google Scholar] | [Crossref]
- Beatrice S, Marco P, Elisa R. Virulence genes and multidrug resistance in avian Ecoli. Vet Microbiol 2025;285:110615. [Google Scholar] | [Crossref]
- Rafiq S, Khan A, Javed H. Antibiotic resistance and virulence profiling in poultry Ecoli. Int J Vet Sci 2024;15(3):112-123. [Google Scholar] | [Crossref]
- Shobrak M, Hassan S, Stiévenart C, El-Deeb B, Gherbawy Y. Prevalence and antibiotic resistance profile of intestinal bacteria isolated from captive adult Houbara bustards (Chlamydotis macqueenii) exposed to natural weather conditions in Saudi Arabia. Glob Veterinaria 2013;10(3):276-284. [Google Scholar] | [Crossref]
- Huff W, Rath N, Balog J. Environmental stressors and colibacillosis in poultry. Poult Sci 2015;94(10):2319-2329. [Google Scholar] | [Crossref]
- El-Gazzar M, Kang S. Stress and immune function in captive birds. J Avian Med Surg 2024;38(2):145-156. [Google Scholar] | [Crossref]
- Gedeno J, Otiang E, Makau D. Risk factors for Ecoliinfections in juvenile poultry. Vet Rec 2022;191(5):205-212. [Google Scholar] | [Crossref]
- Joseph J, Jennings M, Barbieri N, Zhang L, Adhikari P, Ramachandran R. Characterization of avian pathogenic Escherichia coli isolated from broiler breeders with colibacillosis in Mississippi. Poultry 2023;2(1):24-39. [Google Scholar] | [Crossref]
- Shterzer N, Rothschild N, Sbehat Y, Dayan J, Eytan D, Uni Z. Vertical transmission of gut bacteria in commercial chickens is limited. Anim Microbiome 2023;5:27. [Google Scholar] | [Crossref]
- Risalvato J, Sewid A, Eda S, Gerhold R, Wu J. Strategic detection of Escherichia coli in the poultry industry:food safety challenges, One Health approaches, and advances in biosensor technologies. Biosensors 2025;15:419. [Google Scholar] | [Crossref]
- Tilli G, Rossi L, Bianchi A, Conti P. A systematic review on the role of biosecurity to prevent or control colibacillosis in broiler production. Poult Sci 2024;103(7):103642. [Google Scholar] | [Crossref]
- Long JR, Moore SJ, Woodward MJ. Longitudinal study on background lesions in broiler breeder flocks and their progeny, and genomic characterization of Escherichia coli. Vet Res 2022;53:106. [Google Scholar] | [Crossref]
- Rychlik I. Vertical transmission of gut bacteria in commercial chickens is limited. Anim Microbiome 2023;5:35. [Google Scholar] | [Crossref]
- Ibrahim G, Salah-Eldein A, Al-Zaban M, El-Oksh A, Ahmed E, Farid D. Monitoring the genetic variation of some Escherichia coli strains in wild birds and cattle. Onderstepoort J Vet Res 2023;90((1)):a2085. [Google Scholar] | [Crossref]
- Liao M, Wu J, Li Y, Lu X, Tan H, Chen S. Prevalence and persistence of ceftiofur-resistant Escherichia coli in a chicken layer breeding program. Animals 2023;13(1):90. [Google Scholar] | [Crossref]
- Ahmed N, Gulhan T. Determination of antibiotic resistance patterns and genotypes of Escherichia coli isolated from wild birds. Microbiome 2024;12(1):8. [Google Scholar] | [Crossref]
- Osman K, Kappell A, Elhadidy M, ElMougy F, Abd El-Ghany W, Orabi A. Poultry hatcheries as potential reservoirs for antimicrobial-resistant Escherichia coli:a risk to public health and food safety. Sci Rep 2018;8:5859. [Google Scholar] | [Crossref]
- Hosuru subramanya S, Bairy I, Nayak N, Padukone S, Sathian B, Gokhale S. Low rate of gut colonization by extended spectrum β-lactamase producing Enterobacteriaceae in HIV infected persons as compared to healthy individuals in Nepal. PLoS ONE 2019;14(2):e0212042. [Google Scholar] | [Crossref]
- Yang Y, Ashworth A, Willett C, Cook K, Upadhyay A, Owens P. Review of antibiotic resistance, ecology, dissemination, and mitigation in U. S. broiler poultry systems. Front Microbiol 2019;10:2. [Google Scholar] | [Crossref]
- Rousham E, Unicomb L, Islam M. Human, animal and environmental contributors to antibiotic resistance in low-resource settings:integrating behavioural, epidemiological and One Health approaches. Proc R Soc B Biol Sci 2018;285(1876):2018033. [Google Scholar] | [Crossref]
- Falgenhauer L, Imirzalioglu C, Oppong K, Akenten C, Hogan B, Krumkamp R. Detection and characterization of ESBL-producing Escherichia coli from humans and poultry in Ghana. Front Microbiol 2019;9:3358. [Google Scholar] | [Crossref]
- Borzi M, Sordi M, Barbieri R. Synergistic action of virulence factors in avian Ecoli. Vet Microbiol 2018;220:1-9. [Google Scholar] | [Crossref]
- Amer M, Mekky H, Amer A, Fedawy H. Antimicrobial resistance genes in pathogenic Escherichia coli isolated from diseased broiler chickens in Egypt and their relationship with the phenotypic resistance characteristics. Vet World 2018;11((8)). [Google Scholar] | [Crossref]
- Wang J, Tang P, Tan D, Wang L, Zhang S, Qiu Y. The pathogenicity of chicken pathogenic Escherichia coli is associated with the numbers and combination patterns of virulence associated genes. Open J Vet Med 2015;5((12)). [Google Scholar] | [Crossref]
- Sadek D, Rady M, Fedawy H, Rabie N. Molecular epidemiology and sequencing of avian pathogenic Escherichia coli (APEC) in Egypt. Adv Anim Vet Sci 2020;8(5):499-505. [Google Scholar] | [Crossref]
- Collingwood C, Kemmett K, Williams N, Wigley P. Is the concept of avian pathogenic Escherichia coli as a single pathotype fundamentally flawed?. Front Vet Sci 2014;1:5. [Google Scholar] | [Crossref]
- Heidemann Olsen R, Thøfner I, Pors S, Pires dos Santos T, Christensen J. Experimental induced avian Escherichia coli salpingitis:significant impact of strain and host factors on the clinical and pathological outcome. Vet Microbiol 2016;188:59-66. [Google Scholar] | [Crossref]