

ABSTRACT
Background and Aim: Enteric methane emissions from ruminants contribute significantly to greenhouse gas production, prompting research into sustainable feed additives. Macroalgae, rich in bioactive compounds, show promise in modulating rumen fermentation, improving digestibility, and reducing methane output. Indonesia's diverse native macroalgae remain largely unexplored for these purposes, necessitating comparative evaluation to identify promising species for ruminant nutrition. This study aimed to conduct the first comparative in vitro evaluation of rumen fermentation patterns, digestibility characteristics, gas production kinetics, and enteric methane emissions using 14 native Indonesian macroalgae species, including brown (
Materials and Methods: Macroalgae samples were collected from various Indonesian locations, dried, and analyzed for chemical composition (dry matter, ash, crude protein, crude fat, crude fiber, nitrogen-free extract).
Results: Chemical composition varied widely; red algae like Palmaria palmata had high crude protein (22.39 % dry matter), while brown algae like Padina sp. were ash-rich (74.39 % dry matter). Total gas production was highest in B. forbesii (54.75 mL; p < 0.01) and lowest in T. ornata (10.94 mL). Methane emissions and methane per incubated dry matter were lowest in Sargassum sp. (1.87 mM and 3.75 mM/g dry matter; p < 0.01), with Sargassum sp. and C. racemosa reducing methane by 71.86 %. Ammonia levels were similar across species (p > 0.05). Total VFA and propionate were highest in H. durvillei and B. forbesii (p < 0.01), with reduced acetate-to-propionate ratios. IVDMD and IVOMD were highest in H. durvillei (81.72 % and 69.53 %; p < 0.01). Gas kinetics showed B. forbesii with the highest asymptote (201.97 mL; p < 0.01) but slowest rate (0.01 mL/h). Positive correlations existed between crude protein and VFA/ammonia, while crude fiber inversely correlated with gas production and digestibility.
Conclusion:
Keywords: comparative macroalgae, enteric methane mitigation, gas production kinetics,
INTRODUCTION
The increasing global demand for sustainable livestock production has intensified research into alternative feed resources capable of mitigating the environmental footprint of ruminant agriculture, particularly by reducing enteric methane (CH4) emissions. Macroalgae have attracted considerable attention as a promising dietary intervention due to their distinctive biochemical profiles, capacity to modulate rumen fermentation patterns, and documented potential to suppress methanogenesis [1]. Macroalgae exhibit remarkable morphological and functional diversity and are taxonomically classified into three major groups: brown algae (Phaeophyceae, Heterokontophyta), red algae (Rhodophyta), and green algae (Chlorophyta) [2]. Their nutritional composition, including protein, minerals, polysaccharides, lipids, and bioactive secondary metabolites, varies substantially depending on species, geographic origin, season of harvest, and prevailing environmental factors such as light intensity, water temperature, and nutrient availability [3]. When included in ruminant diets, macroalgae can provide essential macro- and micronutrients while simultaneously influencing rumen microbial ecology and fermentation dynamics, often leading to improvements in animal health, nutrient utilization efficiency, and overall productivity [4]. These beneficial effects are primarily attributed to the presence of secondary metabolites—such as phlorotannins, polyphenols, and halogenated compounds (notably bromoform)—which directly inhibit methanogenic archaea, together with shifts in volatile fatty acid (VFA) profiles that enhance propionate production and improve hydrogen sink efficiency [5–7].
More recent investigations have revealed that macroalgae not only lower enteric methane output but also induce significant changes in rumen microbial community structure and gene expression. For example, red seaweeds belonging to the genus
Despite the growing global interest in macroalgae as anti-methanogenic feed additives, several critical knowledge gaps persist, particularly in tropical and biodiverse regions such as Indonesia. Most published studies have focused on a limited number of cosmopolitan or commercially cultivated species (especially
The primary aim of this study was to address the identified research gap by conducting the first comprehensive comparative
MATERIALS AND METHODS
Ethical approval
Rumen fluid containing solid particles was used as the source of microbial inoculum in this study. Rumen fluid was collected from three Ongole crossbreed cattle (
Study duration and location
The
Collection and identification of macroalgae
A total of 14 macroalgae species were evaluated, namely
Table 1. Some of the marine macroalgae species used in this study with sampling site location coordinates.
| No. | Class | Order | Species | GPS coordinates | Sampling location |
|---|---|---|---|---|---|
| 1 | Florideophyceae | Halymeniales | 7.1830° S, 131.7375° E | Maluku (Larat), Indonesia | |
| 2 | Florideophyceae | Gracilariales | 6°01’46” S, 106°26’30” E | Tangerang (Banten), Indonesia | |
| 3 | Ulvophyceae | Bryopsidales | 5.5966° S, 119.4832° E | Makassar (Takalar), Indonesia | |
| 4 | Florideophyceae | Palmariales | 7.1830° S, 131.7375° E | Maluku (Larat), Indonesia |

Figure 1. Sampling locations of
Sample processing and storage
Macroalgae samples were subjected to a two-stage drying process consisting of sun drying for three days followed by oven drying at 55°C for four days. Dried samples were ground using a 0.5-mm disc mill [10]. Sample preparation followed previously described drying and processing procedures [11]. The processed seaweed flour was used for chemical composition analysis and
Taxonomic and phycological identification was conducted for selected macroalgae species, including
Chemical composition analysis of macroalgae
Dry matter, organic matter, ash, crude protein, crude fiber, and ether extract contents were determined using AOAC standard methods [12]. Proximate analysis was conducted in duplicate (n = 2) for each parameter. The
Table 2. Chemical composition (% dry matter) of several macroalgal species used in the study.
| Macroalgae species | % Dry matter | % Moisture | % Ash | % Crude protein | % Crude fat | % Crude fiber | % Nitrogen-free extract (NFE) |
|---|---|---|---|---|---|---|---|
| Brown seaweeds | |||||||
| | 96.88 | 3.22 | 74.39 | 4.46 | 0.54 | 8.71 | 11.91 |
| | 90.22 | 10.85 | 23.50 | 11.91 | 1.31 | 5.34 | 57.94 |
| | 93.11 | 7.40 | 36.82 | 10.55 | 1.46 | 9.21 | 41.96 |
| Green seaweeds | |||||||
| | 94.54 | 5.77 | 43.01 | 5.93 | 1.54 | 25.39 | 24.13 |
| | 92.54 | 8.07 | 43.23 | 11.36 | 1.31 | 5.21 | 38.90 |
| | 87.61 | 14.14 | 25.21 | 14.73 | 0.64 | 8.46 | 50.96 |
| Red seaweeds | |||||||
| | 90.41 | 10.60 | 35.24 | 10.59 | 2.10 | 8.14 | 43.93 |
| | 90.08 | 11.01 | 30.84 | 5.19 | 0.84 | 6.14 | 56.99 |
| | 91.76 | 8.98 | 31.50 | 11.15 | 0.50 | 47.34 | 9.51 |
| | 91.65 | 9.11 | 20.60 | 7.18 | 0.71 | 15.89 | 55.61 |
| | 89.48 | 11.76 | 11.97 | 13.01 | 0.26 | 8.12 | 66.64 |
| | 91.50 | 9.28 | 14.08 | 11.44 | 1.95 | 9.51 | 63.02 |
| | 83.01 | 20.47 | 59.47 | 10.77 | 0.66 | 3.31 | 25.79 |
| | 94.20 | 6.15 | 15.19 | 22.39 | 0.33 | 8.39 | 53.70 |
NFE was calculated as 100 - (% ash + % crude fiber + % crude fat + % crude protein). All values are expressed on a dry matter basis.
In vitro incubation procedure
McDougall buffer solution was preheated to 39°C and flushed with CO2 before mixing with rumen fluid at a ratio of 1:2 (v/v). For each incubation unit, 500 mg of substrate was placed into a 100-mL serum bottle, followed by the addition of 50 mL of buffered rumen fluid. Anaerobic conditions were established by CO2 flushing before sealing the bottles with rubber stoppers and aluminium crimps. Incubations were performed in a water bath at 39°C for 48 or 72 h. After 48 h, samples were centrifuged at 378 ×
Gas production measurement and methane estimation
Cumulative gas production was recorded at 2, 4, 6, 8, 10, 12, 24, 48, and 72 h of incubation using calibrated 50-mL medical syringes (Thermo Fisher Scientific, USA). Bottles were manually shaken after each measurement. Blank bottles containing rumen fluid and buffer without substrate were included for baseline gas correction.
Methane production (CH4 mM) was estimated from VFA profiles using a stoichiometric equation described previously [14]:
CH4 = 0.5C2 + 0.5C4 − 0.25C3 − 0.25C5,
where C2, C3, C4, and C5 represent acetate, propionate, butyrate, and valerate, respectively. Estimated methane production per incubated dry matter (mM/g DM) was calculated by dividing total CH4 by the dry matter weight of the incubated substrate.
Fermentation parameters and in vitro digestibility analysis
Rumen pH was measured immediately after incubation using a calibrated pH meter (BP3001, Trans Instruments). Supernatant and solid fractions were separated after incubation. Supernatants were analyzed for total VFA, SCFA profiles, acetate-to-propionate ratio, and NH3–N concentration, while solid residues were used to determine DMD and OMD.
Total VFA and SCFA concentrations were quantified using GC-MS (Shimadzu-QP2010 SE, Japan) as described previously [15]. SCFA analysis was performed using a MEGA-WAX MS column with helium as the carrier gas at a flow rate of 3 mL/min. Sample preparation, chromatographic conditions, and compound identification followed established protocols [15].
Ammonia nitrogen (NH3–N) concentration was determined spectrophotometrically using the phenol–hypochlorite method described earlier [16]. Absorbance was measured at 630 nm using a Shimadzu UV-1800 portable spectrophotometer.
After
Partitioning factor and microbial protein estimation
PF was calculated as the ratio of dry matter degraded to total gas production and used as an indicator of microbial biomass synthesis efficiency [18]. Microbial protein synthesis was estimated based on OMD using the following equation described previously [19]:
Microbial protein (g/kg organic matter digestibility) = OMD (g) × 19.3 × 6.25
Gas production kinetics
Total gas production kinetics were evaluated using Ørskov’s exponential model described previously [20, 21]:
p = a + b (1 − e-ct),
where p represents cumulative gas production, a is gas from the soluble fraction, b is gas from the insoluble fraction, c is the gas production rate constant, and t is incubation time.
Statistical analysis
Data were analyzed using analysis of variance (ANOVA) in IBM SPSS version 22.0. The experimental design was a completely randomized design comprising 14 macroalgae treatments with five replicates per treatment. The statistical model applied was:
Yij = μ + τi + εij
Where:
Yij= observation from treatment i, replicate j
μ = overall mean
𝜏i = treatment effect
𝜀ij = random error
Differences among treatment means were evaluated using Duncan’s post-hoc test. Statistical analysis followed established procedures described previously [22].
RESULTS AND DISCUSSION
In vitro rumen fermentation profiles of macroalgae
Total gas production differed significantly among the macroalgal species tested.
Table 3.
| Macroalgae species | Total gas production (mL) | Estimated methane production (mM) | Estimated methane production (mM/g incubated DM) | Estimated methane production/total gas production (mM/mL) |
|---|---|---|---|---|
| Brown seaweeds | ||||
| | 17.30 ± 6.92ab | 23.87 ± 6.22cd | 47.72 ± 12.44ef | 1.81 ± 1.36ab |
| | 30.83 ± 9.19abc | 1.87 ± 0.08a | 3.75 ± 0.17a | 0.06 ± 0.03a |
| | 10.94 ± 7.75a | 11.88 ± 7.94abc | 23.76 ± 15.88abc | 1.58 ± 1.30bc |
| Green seaweeds | ||||
| | 54.75 ± 4.84c | 15.32 ± 10.94abc | 30.63 ± 21.87bcdef | 0.50 ± 0.62ab |
| | 19.74 ± 9.53ab | 6.66 ± 3.40ab | 13.31 ± 6.79ab | 0.59 ± 0.72bc |
| | 31.91 ± 15.90abc | 22.63 ± 14.40cd | 45.25 ± 28.79def | 1.43 ± 1.85abc |
| Red seaweeds | ||||
| | 19.36 ± 9.90ab | 19.11 ± 7.76bcd | 38.20 ± 15.52cde | 1.93 ± 2.57abc |
| | 18.74 ± 13.60abc | 14.03 ± 7.87abc | 28.05 ± 15.73abcde | 0.68 ± 0.39abc |
| | 18.50 ± 8.74ab | 11.11 ± 2.74abc | 22.22 ± 5.48abcd | 0.71 ± 0.32ab |
| | 23.89 ± 13.96abc | 11.31 ± 8.29abc | 22.62 ± 16.58abcde | 0.92 ± 0.62c |
| | 35.16 ± 16.91bc | 20.82 ± 8.70bcd | 41.63 ± 17.40cdef | 0.96 ± 1.02abc |
| | 29.23 ± 12.54abc | 21.51 ± 8.06bcd | 43.01 ± 16.12cdef | 0.92 ± 0.62abc |
| | 14.07 ± 6.40ab | 31.80 ± 7.29d | 63.58 ± 14.58f | 3.11 ± 2.54ab |
| | 21.89 ± 9.84ab | 13.46 ± 3.47abc | 26.91 ± 6.93abcde | 0.75 ± 0.40ab |
| p-value | 0.01 | 0.01 | 0.01 | 0.05 |
Means with different superscripts within a column are significantly different (p < 0.01 and p < 0.05). Values are presented as mean ± standard deviation. DM = dry matter.
Means with different superscripts within a column are significantly different (p < 0.01 and p < 0.05). Values are presented as mean ± standard deviation. DM = dry matter.
This pattern indicates that
The efficiency of converting digested dry matter into microbial biomass (rather than gas) was broadly similar across species. This consistency suggests that the primary differences lie in how each macroalga partitions metabolized nutrients, some favoring microbial growth, others directing more toward gas production.
Among all species evaluated,
Overall implications for species selection
These findings emphasize the critical importance of species-specific selection when incorporating macroalgae into ruminant diets. Highly fermentable and digestible species (e.g.,
Careful selection based on these fermentation characteristics allows optimization of macroalgae as sustainable feed components, balancing nutrient supply, microbial efficiency, and environmental benefits (particularly methane reduction) in ruminant production systems.
Methane production and mitigation potential
Methane production (expressed as millimolar concentration, mM) varied substantially among the macroalgal species (p < 0.01; Table 3).
The methane-to-gas production ratio was also strongly affected (p < 0.01). Overall, the highly significant differences (p < 0.01) demonstrated that macroalgae species differ markedly in their fermentation characteristics and methane emission potential.
Among the species tested,
These results highlight the potential of selected macroalgae to serve as targeted functional feed additives, offering a practical approach to enteric methane reduction without severely compromising overall fermentation or nutrient supply in ruminant diets.
Methane inhibition mechanisms and species-specific effects
High methane inhibition by
In contrast,
These species-specific differences highlight the importance of targeted assessments when selecting macroalgae for sustainable ruminant feed formulations and methane mitigation programs.
Variation in estimated methane reduction
Estimated methane production decreased to varying degrees across the macroalgal species evaluated. Distinct differences were observed among treatments, revealing substantial variation in methane suppression potential. Several species achieved significantly lower approximated methane production than others, while some showed only modest reductions. Overall, the extent of methane mitigation was highly species-dependent (Figure 2)
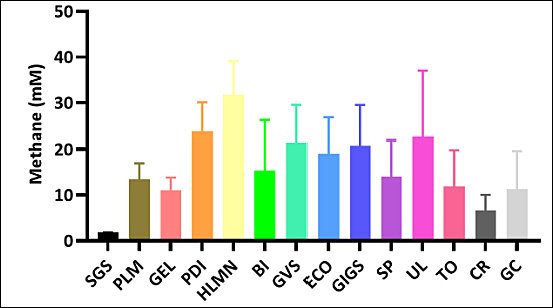
Figure 2. Reduction of estimated methane production (mM) across macroalgae species. Reduction of estimated methane production between macroalgae (mM). SGS =
Rumen pH and fermentation stability
Table 4.
| Macroalgae species | pH | Ammonia (N-NH3) (mg/dL) | Partitioning Factor (mg DMD/mL gas) | Microbial protein (g/kg OMD) |
|---|---|---|---|---|
| Brown seaweeds | ||||
| | 6.98 ± 0.05abc | 55.63 ± 29.24 | 17.30 ± 6.92 | 48.22 ± 35.27abcd |
| | 7.04 ± 0.11c | 56.23 ± 15.67 | 30.83 ± 9.19 | 16.19 ± 3.82a |
| | 6.97 ± 0.07abc | 56.27 ± 16.56 | 10.94 ± 7.75 | 14.30 ± 7.83a |
| Green seaweeds | ||||
| | 6.92 ± 0.10abc | 63.59 ± 23.76 | 54.75 ± 4.84 | 73.89 ± 41.35bcde |
| | 6.83 ± 0.07a | 48.45 ± 18.26 | 19.74 ± 9.53 | 27.94 ± 8.80ab |
| | 6.87 ± 0.06ab | 58.34 ± 19.36 | 31.91 ± 15.90 | 87.70 ± 10.31de |
| Red seaweeds | ||||
| | 6.95 ± 0.08abc | 59.84 ± 20.05 | 19.36 ± 9.86 | 62.63 ± 12.82bcde |
| | 6.95 ± 0.03abc | 41.16 ± 24.44 | 18.75 ± 13.59 | 111.75 ± 30.93e |
| | 7.01 ± 0.06bc | 56.45 ± 19.13 | 18.50 ± 8.74 | 15.90 ± 5.31a |
| | 6.95 ± 0.06abc | 49.92 ± 12.76 | 23.89 ± 13.96 | 11.65 ± 2.99ab |
| | 6.96 ± 0.07abc | 55.20 ± 32.32 | 35.16 ± 16.91 | 62.81 ± 7.71bcde |
| | 7.01 ± 0.11bc | 55.99 ± 24.20 | 29.23 ± 12.54 | 60.87 ± 13.05bcde |
| | 6.94 ± 0.15abc | 75.84 ± 31.21 | 14.07 ± 6.40 | 76.96 ± 20.53cde |
| | 7.01 ± 0.11bc | 54.52 ± 9.02 | 21.89 ± 9.84 | 44.41 ± 8.53abc |
| p-value | <0.05 | 0.068 | 0.171 | 0.01 |
Means with different superscripts within a column are significantly different (p < 0.01 and p < 0.05). Values are presented as mean ± standard deviation. DMD = dry matter digestibility, OMD = organic matter digestibility.
Certain macroalgae may exert mild buffering or acidifying effects on the rumen environment. However, rumen fermentation parameters remained near neutrality across all treatments (Table 4), indicating that macroalgae inclusion did not cause harmful ruminal conditions.
Ammonia concentrations and microbial protein synthesis
Ammonia (NH3) concentrations showed no significant differences among macroalgal treatments (p > 0.05; Table 4). Nevertheless,
The elevated ammonia in
Ammonia concentrations across all species were sufficient to support microbial growth, despite some numerical variation. Microbial protein synthesis differed significantly among treatments, with higher production observed in
Partitioning factor and microbial efficiency
The partitioning factor (PF; Table 4; p > 0.05) showed wide variation among the macroalgal substrates. PF is calculated as the ratio of truly degraded organic matter (mg) to gas produced (mL). Higher PF values indicate greater fermentation efficiency, with a larger proportion of metabolized dry matter retained as microbial biomass rather than lost as gas.
Microbial protein production differed significantly among treatments (p < 0.05). Certain macroalgae strongly supported high microbial protein synthesis; for example, one sample yielded 111.75 g of microbial protein per kg of organic matter degraded (OMD), demonstrating excellent microbial efficiency.
Ammonia dynamics and protein fractions
Decreases in ammonia concentration generally reflect increased incorporation of ammonia-nitrogen (NH3-N) into microbial protein by rumen microorganisms. This process can be constrained by reduced proteolysis or insufficient fermentable carbohydrate availability [25].
Different macroalgae species contain varying proportions of rumen-degradable protein (RDP; protein broken down in the rumen to supply ammonia and peptides for microbial growth) and rumen-undegradable protein (RUP; bypass protein that escapes rumen fermentation and is digested in the small intestine for direct benefit to the host animal) [26, 27]. Further detailed characterization of RDP and RUP fractions in macroalgae is warranted. Optimizing RUP content could significantly improve feed efficiency, body weight gain, and milk production in ruminants. Secondary metabolites such as tannins, saponins, and flavonoids are known to influence the RDP/RUP balance by binding proteins or modulating microbial activity.
High NH3-N concentrations typically indicate poor rumen ammonia absorption, excessive deamination of amino acids, or slow microbial uptake and metabolism of ammonia [28]. In this study, ammonia levels remained adequate to sustain microbial growth across all species, despite some numerical variation.
Implications for ruminant nutrition and feed formulation
PF and microbial protein synthesis are critical indicators of how effectively rumen microorganisms utilize feed substrates during fermentation. Higher PF values signify better nutrient partitioning toward microbial biomass production, thereby enhancing the supply of microbial protein to the host ruminant and improving overall nitrogen efficiency.
The results demonstrate that macroalgae differ markedly in their capacity to provide fermentable energy and available nitrogen to support rumen microbial growth. Some species promote high microbial protein synthesis and efficient substrate utilization, while others are less effective. Selective inclusion of macroalgae with favorable PF and high microbial protein yields can therefore enhance rumen microbial efficiency, increase protein delivery to the animal, and contribute to more sustainable and productive ruminant feeding systems. It is imperative to exercise caution when selecting species in order to optimize these advantages.
These findings reinforce the value of macroalgae as a diverse resource in ruminant nutrition, where strategic use of high-PF, protein-efficient species can optimize microbial protein supply and nitrogen utilization while minimizing environmental impacts.
VFA production and fermentation pathways
Decreased VFA production in
In contrast,
Table 5.
| Macroalgae species | Total VFA (mM) | Acetate (mM) | Propionate (mM) | Butyrate (mM) | Valerate (mM) | iso-Butyrate (mM) | iso-Valerate (mM) | A/P |
|---|---|---|---|---|---|---|---|---|
| Brown seaweeds | ||||||||
| | 99.15 ± 23.49ab | 55.16 ± 14.28ab | 28.49 ± 6.36abc | 7.09 ± 1.34ab | 0.95 ± 0.18b | 2.81 ± 0.62abc | 4.66 ± 0.96ab | 1.92 ± 0.12cd |
| | 45.08 ± 24.22a | 23.98 ± 12.51a | 13.03 ± 7.33ab | 3.40 ± 2.01ab | 0.41 ± 0.31ab | 1.67 ± 0.79abc | 2.59 ± 1.32ab | 1.87 ± 0.08bcd |
| | 80.31 ± 73.24ab | 46.56 ± 43.57ab | 22.21 ± 20.05abc | 5.28 ± 5.05ab | 1.05 ± 0.70ab | 2.19 ± 1.44abc | 3.44 ± 2.53ab | 2.02 ± 0.16cd |
| Green seaweeds | ||||||||
| | 95.91 ± 66.76ab | 43.66 ± 30.86ab | 38.01 ± 26.36c | 6.39 ± 4.57ab | 1.36 ± 0.19b | 2.68 ± 1.45abc | 4.36 ± 2.80ab | 1.12 ± 0.10a |
| | 30.31 ± 17.09a | 15.84 ± 7.77a | 9.24 ± 5.24a | 2.17 ± 1.72a | 0.27 ± 0.10a | 1.15 ± 0.94a | 1.75 ± 1.46a | 1.83 ± 0.29bcd |
| | 86.53 ± 59.94ab | 54.04 ± 34.11ab | 29.28 ± 16.01abc | 6.23 ± 3.08ab | 0.90 ± 0.42b | 2.74 ± 1.20abc | 4.08 ± 1.88ab | 1.81 ± 0.19bcd |
| Red seaweeds | ||||||||
| | 86.27 ± 31.07ab | 45.24 ± 18.36ab | 26.11 ± 9.13abc | 6.35 ± 1.80ab | 0.88 ± 0.35ab | 2.85 ± 0.55bc | 4.84 ± 1.12b | 1.70 ± 0.14bc |
| | 61.77 ± 33.30ab | 32.87 ± 18.20ab | 18.20 ± 9.74abc | 4.49 ± 2.66ab | 0.66 ± 0.36ab | 2.24 ± 0.86abc | 3.44 ± 1.74ab | 1.81 ± 0.17bcd |
| | 47.95 ± 11.23a | 25.53 ± 6.33a | 13.51 ± 3.42ab | 3.67 ± 0.83ab | 0.50 ± 0.09ab | 1.73 ± 0.31abc | 3.01 ± 0.45ab | 1.89 ± 0.07cd |
| | 51.87 ± 28.17ab | 27.66 ± 17.24a | 16.35 ± 7.69abc | 3.45 ± 2.32ab | 0.65 ± 0.49ab | 1.56 ± 0.62ab | 2.21 ± 1.30ab | 1.72 ± 0.58bcd |
| | 97.85 ± 35.65ab | 50.44 ± 20.63ab | 31.66 ± 10.60abc | 7.45 ± 2.26b | 0.96 ± 0.38b | 2.73 ± 0.72abc | 4.62 ± 1.33ab | 1.56 ± 0.14b |
| | 95.60 ± 34.62ab | 50.87 ± 18.63ab | 29.46 ± 10.45abc | 7.15 ± 3.04ab | 0.84 ± 0.45ab | 2.75 ± 0.82abc | 4.52 ± 1.52ab | 1.72 ± 0.13bcd |
| | 127.51 ± 24.25b | 73.85 ± 16.63b | 36.13 ± 6.43bc | 8.11 ± 1.24b | 1.02 ± 0.21b | 3.30 ± 0.32c | 5.09 ± 0.57b | 2.03 ± 0.16d |
| | 61.99 ± 15.02ab | 31.65 ± 8.30a | 18.39 ± 4.46abc | 4.78 ± 1.08ab | 0.71 ± 0.25ab | 2.45 ± 0.35abc | 4.02 ± 0.85ab | 1.71 ± 0.10bcd |
| p-value | 0.01 | 0.012 | 0.012 | 0.014 | 0.057 | 0.012 | 0.015 | 0.012 |
Means with different superscripts within a column are significantly different (
Species comparisons in VFA profiles
These results highlight distinct fermentation profiles among the macroalgae, with
VFA profiles and implications for methane mitigation
Methane mitigation in
The A/P ratio varied significantly among species (p = 0.012).
Total VFA concentration was highest in
These findings align with previous research on brown macroalgae. For example, three brown species (
NSP and hydrogen sink manipulation
Characterizing the fermentability of individual non-starch polysaccharide components is essential when incorporating macroalgae into ruminant diets. Intact macroalgae rich in non-starch polysaccharides may function as wholesome, fiber-based feed ingredients [30].
Rumen methane production arises from syntrophic interactions between H2-producing microbes and H2-oxidizing methanogens [31]. Manipulating the hydrogen sink is a key strategy for reducing CH4 emissions. Becker
Overall, these VFA profiles reinforce the potential of selected macroalgae, particularly those with favorable propionate promotion or selective inhibition (e.g.,
Correlations between fermentation end-products and methane production
Figure 3 illustrates the correlations between estimated methane production, TVFA, and individual SCFA components. Estimated methane production showed clear associations with both TVFA and specific SCFA fractions. Negative correlations were observed between methane production and propionate concentration, whereas positive correlations existed between methane production and acetate concentration. These relationships confirm consistent links between the distribution of fermentation end-products and methano-genesis: acetate-dominant pathways promote higher methane output, while propionate-promoting pathways divert hydrogen away from methanogens, reducing CH4 formation.
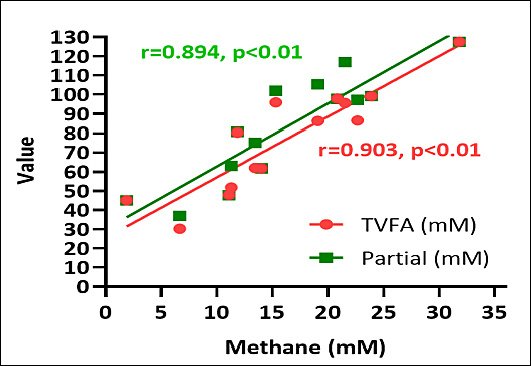
Figure 3. Correlation between estimated methane production (mM), total volatile fatty acid (TVFA), and individual short-chain fatty acid components (mM).
Variations in A/P ratios across species reflect changes in hydrogen utilization directly linked to methane generation. Lower A/P ratios, as observed in
Species-specific VFA profiles and fermentation patterns
Elevated TVFA concentrations in
The relative proportions of individual SCFAs (acetate, propionate, butyrate) varied considerably among species (Figure 4), with some macroalgae favoring acetate- or butyrate-dominant patterns and others promoting elevated propionate. TVFA production showed substantial variability across treatments (Figure 5), confirming species-dependent differences in fermentation intensity.
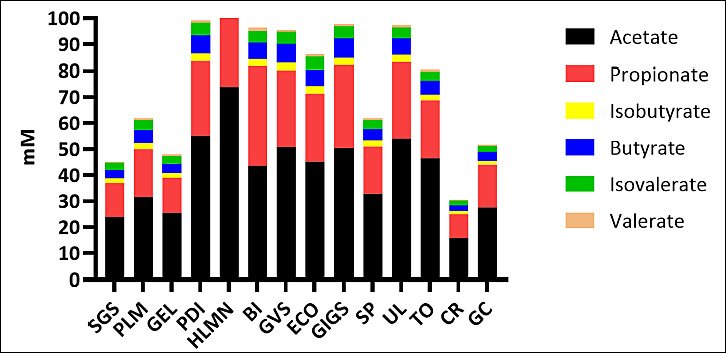
Figure 4. Comparative profile of partial volatile fatty acids (short-chain fatty acids) (mM) across macroalgae species. SGS = Sargassum sp., PLM =
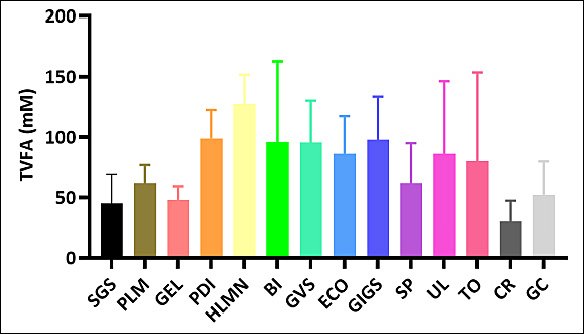
Figure 5. Comparison of total volatile fatty acid (TVFA) profile (mM) across macroalgae species. SGS =
Mechanistic insights and implications
Species with acetate-dominant fermentation patterns generally produced more methane, reinforcing the strong stoichiometric relationship between VFA profiles and methanogenesis. Differences in branched-chain VFA and ammonia concentrations further indicate variation in protein degradation rates and microbial growth efficiency, consistent with the observed disparities in microbial protein synthesis.
Differences in ruminal degradability among macroalgae are likely attributable to variations in cell wall polysaccharides [34]. Previous studies [4, 34] have reported CH4 reductions without significant losses in digestibility [4], aligning with the 24 h
Overall, these VFA profiles, correlations, and species-specific patterns emphasize that macroalgae exert distinct effects on rumen fermentation pathways. Targeted selection of species that promote propionate production (e.g.,
Digestibility and nutrient availability across macroalgal species
Reduced propionate production may be attributed to the inhibition of many key succinate-utilizing bacteria following
Table 6.
| Macroalgae species | IVDMD (%) | IVOMD (%) |
|---|---|---|
| Brown seaweeds | ||
| | 31.60 ± 12.49ab | 39.98 ± 29.24abcd |
| | 26.98 ± 2.31a | 13.42 ± 3.17ab |
| | 24.69 ± 6.08a | 11.85 ± 6.49a |
| Green seaweeds | ||
| | 52.00 ± 4.57cd | 46.03 ± 4.57cde |
| | 47.07 ± 4.11abc | 23.17 ± 7.30abc |
| | 77.93 ± 7.94de | 72.70 ± 8.55cd |
| Red seaweeds | ||
| | 60.21 ± 9.70cd | 51.92 ± 10.63cd |
| | 79.85 ± 6.40e | 74.60 ± 9.63e |
| | 21.78 ± 5.15a | 13.19 ± 4.40ab |
| | 20.85 ± 1.31ab | 9.66 ± 2.48abc |
| | 53.51 ± 5.30bc | 52.07 ± 6.39cd |
| | 52.90 ± 10.55bc | 50.46 ± 10.82bcd |
| | 81.72 ± 11.67de | 69.53 ± 19.54de |
| | 44.19 ± 6.64abc | 36.82 ± 7.07abcd |
| p-value | 0.011 | 0.01 |
Means with different superscripts within a column are significantly different (p < 0.01 and p < 0.05). Values are presented as mean ± standard deviation. IVDMD =
The highest IVDMD and IVOMD were observed in
Red and green macroalgae, including
In other studies,
Digestibility vs. methane mitigation trade-offs
A key finding is the divergence between species with high digestibility and those with strong methane-suppressing potential. Red macroalgae such as
Chemical composition correlations and methane production
Nonetheless, the conditions that enhanced digestibility also facilitated methanogenesis.
The relationship between estimated methane production and macroalgal chemical composition is depicted in Figure 6. Estimated methane production exhibited a negative correlation with crude protein content and a positive correlation with crude fiber content. Macroalgae with elevated crude protein levels generally corresponded with reduced estimated methane production, whereas those with higher crude fiber content were associated with increased methane emissions.
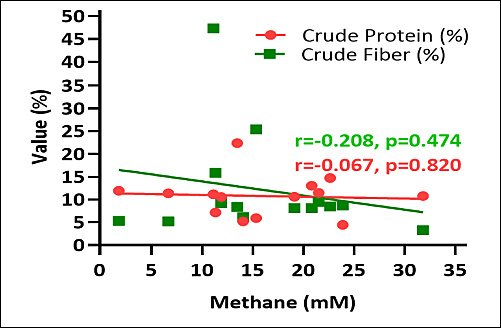
Figure 6. Correlation between estimated methane production (mM), crude protein (%), and crude fiber (%) of various macroalgae species.
Crude fiber content inversely correlated with both TVFA and individual SCFA concentrations. Elevated crude fiber levels were associated with decreased overall VFA production across the examined macroalgae species (Figure 7). Conversely, positive correlations were observed between crude protein content and TVFA and individual SCFA concentrations. Macroalgae with elevated crude protein levels correlated with enhanced TVFA production and increased individual SCFA concentrations. These findings demonstrate a consistent relationship between protein content and fermentation end-product generation (Figure 8).
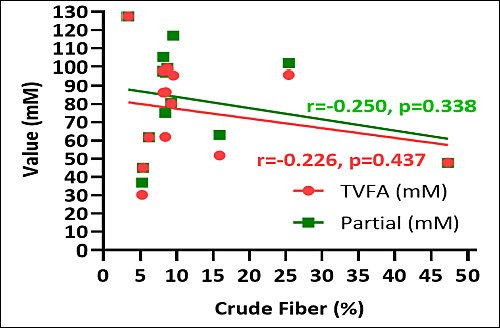
Figure 7. Correlation between total volatile fatty acid (TVFA) (mM), partial volatile fatty acid (short-chain fatty acid) (mM), and crude fiber (%) of various macroalgae species.
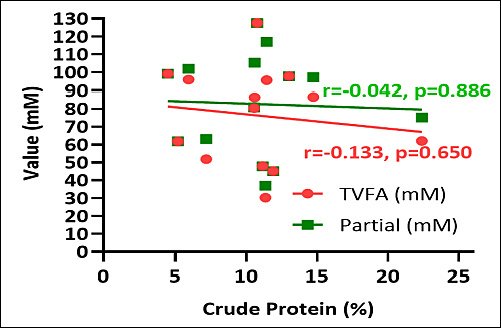
Figure 8. Correlation between total volatile fatty acid (TVFA) (mM), partial volatile fatty acid (short-chain fatty acid) (mM), and crude protein (%) of various macroalgae species.
Factors contributing to interspecies variability
The substantial variability among macroalgae species can be attributed to several key factors. First, chemical composition differed widely, particularly in ash, crude fiber, and non-fiber extract contents, which directly influence rumen degradability and fermentation kinetics. High ash content, common in certain brown and red macroalgae, may dilute fermentable organic matter and limit microbial nutrient access. Second, cell wall structure and polysaccharide composition vary significantly among brown, green, and red macroalgae. Brown species are rich in alginates and fucoidans, red species contain agar and carrageenan, and green species are dominated by cellulose-based polysaccharides. These structural differences affect degradation rates and microbial colonization in the rumen [42]. Third, bioactive compounds, particularly in brown macroalgae such as
Phytochemical content and bioactive potential
Flavonoid content ranged from 0.08 to 1.50 mg quercetin/g, with
Polyunsaturated fatty acids (PUFAs) and methane mitigation
The ability of macroalgal PUFAs to reduce enteric methane emissions during
Bioactive compounds and mechanisms of methane suppression
Although the fatty acid profile and phenolic content were not directly measured in the present study, the observed lower cumulative gas production and methane-related responses, particularly in the
PUFAs, abundant in certain brown macroalgae including
These findings align with literature on brown macroalgae, where secondary metabolites drive enteric methane mitigation. The antimethanogenic effects in
Kinetics of gas production
No statistically significant differences in gas production were observed between 10 and 12 h of incubation (p = 0.067). These early periods likely reflect the latency phase of microbial fermentation, during which gas differences remain minimal. Gas production from
The highest asymptotic gas production (a + b) was recorded in
Table 7.
| Macroalgae Species | Gas10 (mL/g DM) | Gas12 (mL/g DM) | Gas24 (mL/g DM) | Gas36 (mL/g DM) | Gas48 (mL/g DM) | Gas72 (mL/g DM) |
|---|---|---|---|---|---|---|
| Brown seaweeds | ||||||
| | 6.79 ± 3.96ab | 6.79 ± 3.96ab | 17.60 ± 5.94ab | 28.39 ± 6.46abc | 33.79 ± 6.02ab | 41.39 ± 6.23abc |
| | 11.20 ± 9.91ab | 11.20 ± 9.91ab | 33.59 ± 10.40ab | 53.99 ± 11.60fg | 62.99 ± 11.40f | 77.79 ± 13.56ef |
| | 3.20 ± 3.83a | 3.20 ± 3.83a | 10.60 ± 5.85a | 16.79 ± 6.61a | 21.59 ± 6.80a | 26.39 ± 7.89a |
| Green seaweeds | ||||||
| | 12.79 ± 5.45b | 12.79 ± 5.45b | 32.59 ± 8.79b | 60.99 ± 10.08g | 84.59 ± 7.58g | 107.18 ± 9.85g |
| | 7.79 ± 4.87ab | 7.79 ± 4.87ab | 20.19 ± 7.22ab | 31.79 ± 7.32abcd | 38.99 ± 8.18abc | 46.59 ± 9.23abc |
| | 9.99 ± 6.28ab | 9.99 ± 6.28ab | 30.99 ± 10.63ab | 51.59 ± 9.63efg | 60.38 ± 8.56ef | 75.78 ± 9.40ef |
| Red seaweeds | ||||||
| | 9.60 ± 3.91ab | 9.60 ± 3.91ab | 22.40 ± 7.34ab | 33.39 ± 7.13abcde | 39.99 ± 8.25bcd | 47.79 ± 9.20bc |
| | 13.60 ± 5.77b | 13.60 ± 5.77b | 31.79 ± 8.04b | 46.99 ± 10.95cdefg | 53.78 ± 11.52cdef | 61.78 ± 13.18cde |
| | 8.39 ± 5.81ab | 8.39 ± 5.81ab | 19.39 ± 7.43ab | 29.39 ± 8.53abc | 35.79 ± 7.98abc | 42.98 ± 9.77abc |
| | 6.80 ± 9.31ab | 6.80 ± 9.31ab | 21.40 ± 14.64ab | 36.60 ± 16.25bcdef | 44.39 ± 16.90bcde | 55.19 ± 17.78cd |
| | 11.79 ± 7.16ab | 11.79 ± 7.16ab | 38.19 ± 12.32ab | 59.99 ± 12.31g | 70.19 ± 11.48fg | 83.18 ± 12.07f |
| | 10.59 ± 4.39ab | 10.59 ± 4.39ab | 29.80 ± 8.23ab | 48.19 ± 8.44defg | 57.79 ± 8.76def | 70.59 ± 9.29def |
| | 6.79 ± 3.77ab | 6.79 ± 3.77ab | 15.39 ± 4.56ab | 23.39 ± 4.67ab | 28.79 ± 4.60ab | 33.99 ± 4.69ab |
| | 11.99 ± 8.18ab | 11.99 ± 8.18ab | 25.39 ± 11.73ab | 35.99 ± 11.93bcdef | 41.99 ± 12.38bcd | 51.98 ± 14.62bcd |
| p-value | 0.067 | 0.067 | 0.067 | 0.01 | 0.011 | 0.01 |
Means with different superscripts within a column are significantly different (p < 0.01 and p < 0.05). Values are presented as mean ± standard deviation. DM = Dry matter. Gas production values are expressed as mL per g of incubated dry matter (DM).
In this experiment,
In contrast,
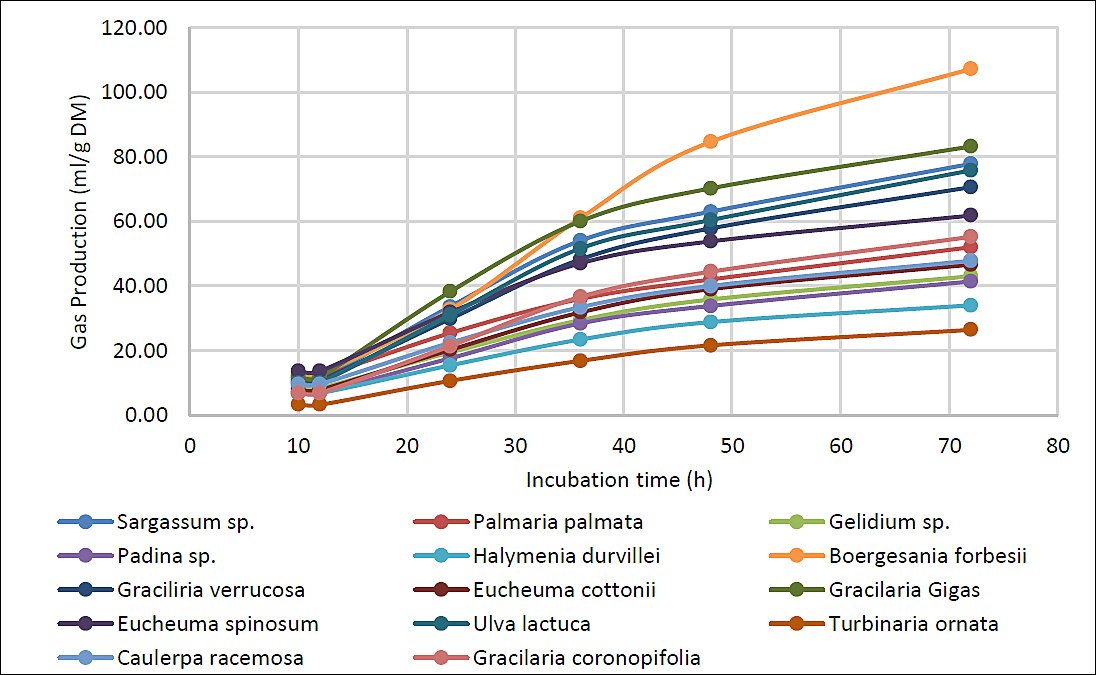
Figure 9. Cumulative gas production (mL/g DM) from the experimental macroalgae diets during the 72-h incubation period.
High-yield but slower-fermenting species like
The asymptote (a + b), rate constant (c), and cumulative gas production decreased as macroalgae inclusion level increased in the substrate, consistent with literature showing general declines in cumulative gas production (CGP) and fermentation with higher inclusion rates. Kinley
Macroalgae species significantly influenced both total gas produced and production rate (p < 0.05 for a + b and c).
Table 8. Gas production parameters derived from varied macroalgae diets over 72 h of incubation.
| Macroalgae Species | a + b (mL) | c (mL/h) |
|---|---|---|
| Brown seaweeds | ||
| | 55.35 ± 8.56abc | 0.02 ± 0.008abc |
| | 97.37 ± 23.14ef | 0.03 ± 0.007bc |
| | 36.12 ± 10.58a | 0.02 ± 0.009abc |
| Green seaweeds | ||
| | 201.97 ± 32.92g | 0.01 ± 0.002a |
| | 59.99 ± 8.68abc | 0.03 ± 0.004bc |
| | 100.30 ± 20.31f | 0.03 ± 0.009bc |
| Red seaweeds | ||
| | 58.39 ± 9.47abc | 0.02 ± 0.002ab |
| | 69.65 ± 14.95bcd | 0.03 ± 0.002bc |
| | 55.14 ± 11.31abc | 0.02 ± 0.006ab |
| | 67.77 ± 6.68cde | 0.02 ± 0.000ab |
| | 97.04 ± 9.39ef | 0.03 ± 0.006c |
| | 90.02 ± 14.82def | 0.02 ± 0.01bc |
| | 44.29 ± 8.83ab | 0.02 ± 0.008abc |
| | 56.98 ± 9.95bc | 0.02 ± 0.005abc |
| p-value | 0.011 | 0.01 |
Means with different superscripts within a column are significantly different (p < 0.01 and p < 0.05). Values are presented as mean ± standard deviation. a + b = Asymptotic (potential) gas production (mL); c = Gas production rate constant (mL/h).
Limitations and practical implications
Despite consistent trends in gas production kinetics, this study was conducted under
Macroalgae should not be considered a uniform ruminant feed resource. Highly digestible species such as
The positive correlation between crude protein content and TVFA concentration/ammonia production aligns with prior research on protein-rich macroalgae and unconventional ruminant feeds [51]. Conversely, the inverse relationship between crude fiber content and total gas production, VFA concentration, and digestibility is consistent with the inhibitory effects of structural carbohydrates on rumen fermentation, as observed in other macroalgae studies. If it is compared to other roughage or forage that the most significant gas production of hay was observed during the 21-day harvesting interval, as well as IVDMD, which contributed to a higher potential for degradation. This occurred as a result of the substantial quantity of soluble material, as the soluble fractions in feed are responsible for the production of volatile fatty acids, which are the primary source of energy for ruminants. The converse condition was observed during the extended cutting intervals; this can be attributed to the degradability of the NDF level, ADF, and lignin contents [52].
A previous study [53], which was about cassava tops (CT) and cassava pulp (CS), there were some nutrient parameters that were different, which may be attributed to the effects of certain compounds in CT and CS on the fermentation process or the varying levels of additive source supplementation. However, those nutrient contents can be utilized as animal feedstuffs to enhance animal productivity.
The gas parameters in the equation account for 74% of the variability in the dry matter effective degradability of the forages, as indicated by the coefficient of determination of 0.740 in the predictor model for the Dry Matter Effective degradability. There was a reduced coefficient of determination for the predictor model for organic matter effective degradability, which was 0.659. The coefficient of determination for effective degradability of CP also showed correlations with cumulative gas productions was the highest, at 0.813. This is a rationale for the conclusion that the majority of the gas generated during fermentation in the rumen is a result of the rumen microbes’ degradation of plant cell wall components. Conversely, the coefficient of determination for the CP ED predictor was only 0.500 [54].
These results suggest that targeted integration of highly digestible and methane-suppressing macroalgae species could provide a balanced strategy for improving feed efficiency and reducing environmental impact in tropical ruminant systems. Further
CONCLUSION
This study provides the first comprehensive
These findings highlight macroalgae as viable, sustainable alternatives to conventional forages in tropical ruminant systems, where forage scarcity is common. High-digestibility species (
The study’s robustness lies in its comparative design across a diverse set of 14 native species, integrating multiple parameters (fermentation, digestibility, kinetics, methane) using standardized
As an
In conclusion, native Indonesian macroalgae represent a promising, underutilized resource for ruminant nutrition, with species like
DATA AVAILABILITY
The data supporting the findings of this study are available upon reasonable request from the corresponding author.
AUTHORS’ CONTRIBUTIONS
DSW: Investigation, validation, data curation, formal analysis, and writing of the original draft. KGW: Conceptualization, supervision, and writing of the review and editing. RR: Conceptualization, supervision, and writing and review of the manuscript. GG: Validation, data analysis, and writing of the manuscript and editing. APS: Validation and writing of the manuscript and editing. MS: Validation and writing of the manuscript and editing. RF and AF: Executed the experiment, performed laboratory analysis, and analyzed the data. DP: Validation and writing, review, and editing of the manuscript. WP: Data analysis and writing, review, and editing of the manuscript. GKA: Experiment execution, laboratory analysis, data analysis, and editing. YW: Validation and writing, review, and editing of the manuscript. AJ: Conceptualization, supervision, methodology, and writing, review, and editing of the manuscript. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors acknowledge the financial support provided by the Research Organization for Agriculture and Food, National Research and Innovation Agency of the Republic of Indonesia (Grant No. 6/III.11/HK/2024) and the RIIM LPDP-BRIN numbers B-803/7.5/FR//2022 and B-173/III.5/PR.03.08/6/2022. The authors express their sincere gratitude to Nurjannah (Department of Fisheries Product Technology, Faculty of Fisheries and Marine Science, IPB University, Indonesia). The authors would like to thank M. Gazali (Department of Marine Science, Faculty of Fisheries and Marine Science Teuku Umar University, West Aceh, Indonesia), Lideman (Research Center for Fishery, National Research and Innovation Agency (BRIN), Indonesia), and Dedi Noviendri (Research Center for Medicinal Raw Materials and Traditional Medicines -National Research and Innovation Agency (BRIN), Indonesia) for their support in sample collection and preparation. We acknowledge the invaluable contributions and critical critiques from several colleagues, which greatly enhanced the study’s design and interpretation.
REFERENCES
- Wasson DE, Stefenoni H, Cueva SF. Screening macroalgae for mitigation of enteric methane
in vitro . Sci Rep 2023;13(1):1-15. [Google Scholar] | [Crossref] - Cardozo KHM, Guaratini T, Barros MP. Metabolites from algae with economical impact. Comp Biochem Physiol C Toxicol Pharmacol 2007;146((1-2)):60-78. [Google Scholar] | [Crossref]
- Michalak I, Tiwari R, Dhawan M. Antioxidant effects of seaweeds and their active compounds on animal health and production-a review. Vet Q 2022;42(1):48-67. [Google Scholar] | [Crossref]
- Machado L, Magnusson M, Paul NA. Effects of marine and freshwater macroalgae on
in vitro total gas and methane production. PLoS One 2014. [Google Scholar] | [Crossref] - Nunes HPB, Maduro Dias CSAM, Álvaro NV, Borba AES. Evaluation of two species of macroalgae from Azores Sea as potential reducers of ruminal methane production:
in vitro ruminal assay. Animals 2024. [Google Scholar] | [Crossref] - Roque-Jiménez A, Ivan J, Chay-Canul AJ. Evaluation of three marine algae on degradability,
in vitro gas production, and CH₄and CO₂emissions by ruminants 2022. [Google Scholar] | [Crossref] - Vargas JE, Andrés S, López-Ferreras L. Dietary supplemental plant oils reduce methanogenesis from anaerobic microbial fermentation in the rumen. Sci Rep 2020;10(1):1-9. [Google Scholar] | [Crossref]
- Zhang P, Roque B, Romero P. Red seaweed supplementation suppresses methanogenesis in the rumen, revealing potentially advantageous traits among hydrogenotrophic bacteria. Microbiome 2025. [Google Scholar] | [Crossref]
- Li S, Sun Y, Guo T.
Sargassum mcclurei mitigating methane emissions and affecting rumen microbial community inin vitro rumen fermentation. Animals 2024. [Google Scholar] | [Crossref] - Paga A, Agus A, Kustantinah, Budisatria IGS. Effect of different drying methods on the mineral content of seaweed
Sargassum sp. Livest Res Rural Dev 2021;33(3):219-223. [Google Scholar] | [Crossref] - Jayanegara A, Wina E, Soliva CR. Dependence of forage quality and methanogenic potential of tropical plants on their phenolic fractions as determined by principal component analysis. Anim Feed Sci Technol 2011;163((2-4)):231-243. [Google Scholar] | [Crossref]
- Official Methods of Analysis of AOAC International 2005. [Google Scholar] | [Crossref]
- Theodorou MK, Williams BA, Dhanoa MS. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim Feed Sci Technol 1994;48((3-4)):185-197. [Google Scholar] | [Crossref]
- Hegarty RS, Nolan JV. Chapter 4 Estimation of ruminal methane production from measurement of volatile fatty acid production 2007:69-70. [Google Scholar] | [Crossref]
- Sarwono KA, Rohmatussolihat R, Watman M, Ratnakomala S. Characteristics of fresh rice straw silage quality prepared with addition of lactic acid bacteria and crude cellulase 2022;7((May)):481-499. [Google Scholar] | [Crossref]
- Detmann E, Filho SCV, Pina DS. Arq.bras 2013;3:1752-1758. [Google Scholar] | [Crossref]
- Tilley JMA, Terry RA. A two-stage technique for the
in vitro digestion of forage crops. J Br Grassl Soc 1963. [Google Scholar] | [Crossref] - Blummel BM.
In vitro gas production:a technique revisited 1997;77:24-34. [Google Scholar] | [Crossref] - Czerkawski JW, Czerkawski JW. Chapter 8 - Synthetic Processes in the Rumen. Amsterdam: Pergamon; 1986. p. 127-150. [Google Scholar]
- Orskov ER, Mcdonald I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J Agric Sci 1979;92(2):499-503. [Google Scholar] | [Crossref]
- Jayanegara A, Sofyan A, Makkar HPS, Becker K. Kinetika produksi gas, kecernaan bahan organik dan produksi gas metana
in vitro pada hay dan jerami yang disuplementasi hijauan mengandung tanin. Media Peternakan 2009;32(2):120-129. [Google Scholar] | [Crossref] - Steel RG, Torrie JH. Principles and Procedures of Statistics:A Biometrical Approach. New York: McGraw-Hill; 1981. [Google Scholar]
- Melgar A, Welter KC, Nedelkov K. Dose-response effect of 3-nitrooxypropanol on enteric methane emissions in dairy cows 2020:6145-6156. [Google Scholar] | [Crossref]
- A XL, A HCN, B RDK.
Asparagopsis taxiformis decreases enteric methane production from sheep 2016. [Google Scholar] | [Crossref] - Wang Y, Xu Z, Bach SJ, McAllister TA. Effects of phlorotannins from
Ascophyllum nodosum (brown seaweed) onin vitro ruminal digestion of mixed forage or barley grain. Anim Feed Sci Technol 2008;145((1-4)):375-395. [Google Scholar] | [Crossref] - Tedeschi LO, Fox DG, Fonseca MA, Francis L. Revista Brasileira de Zootecnia Invited Review Models of protein and amino acid requirements for cattle 2015;44(3):109-132. [Google Scholar] | [Crossref]
- Sharif M, Qamar H, Wahid AA. Effect of rumen degradable protein concentrations on nutrient digestibility, growth performance and blood metabolites in Beetal kids 2019:249-253. [Google Scholar] | [Crossref]
- Tan P, Liu H, Zhao J. Amino acids metabolism by rumen microorganisms:nutrition and ecology strategies to reduce nitrogen emissions from the inside to the outside. Sci Total Environ 2021;800:149596. [Google Scholar] | [Crossref]
- Sanders J, Xie Y, Gazzola D. A new paraprobiotic-based treatment for control of
Haemonchus contortus in sheep. Int J Parasitol Drugs Drug Resist 2020;14((September)):230-236. [Google Scholar] | [Crossref] - Metzler-Zebeli BU, Hooda S, Pieper R. Nonstarch polysaccharides modulate bacterial microbiota, pathways for butyrate production, and abundance of pathogenic
Escherichia coli in the pig gastrointestinal tract. Appl Environ Microbiol 2010;76(11):3692-3701. [Google Scholar] | [Crossref] - Kobayashi Y. Abatement of methane production from ruminants:trends in the manipulation of rumen fermentation. Asian-Australas J Anim Sci 2010;23(3):410-416. [Google Scholar] | [Crossref]
- Becker PM, van Wikselaar PG, Franssen MCR. Evidence for a hydrogen-sink mechanism of (+)-catechin-mediated emission reduction of the ruminant greenhouse gas methane. Metabolomics 2014;10(2):179-189. [Google Scholar] | [Crossref]
- Min BR, Parker D, Brauer D. The role of seaweed as a potential dietary supplementation for enteric methane mitigation in ruminants:challenges and opportunities. Anim Nutr 2021;7(4):1371-1387. [Google Scholar] | [Crossref]
- Belghit I, Rasinger JD, Heesch S. In-depth metabolic profiling of marine macroalgae confirms strong biochemical differences between brown, red and green algae. Algal Res 2017;26((April)):240-249. [Google Scholar] | [Crossref]
- O'Hara E, Terry SA, Moote P. Comparative analysis of macroalgae supplementation on the rumen microbial community:
Asparagopsis taxiformis inhibits major ruminal methanogenic, fibrolytic, and volatile fatty acid-producing microbesin vitro . Front Microbiol 2023;14((April)):1-16. [Google Scholar] | [Crossref] - Herremans S, Vanwindekens F, Decruyenaere V. Effect of dietary tannins on milk yield and composition, nitrogen partitioning and nitrogen use efficiency of lactating dairy cows:a meta-analysis 2020:1-10. [Google Scholar] | [Crossref]
- Carro MD, Rodela MY, Weisbjerg MR, Lind V.
In vitro ruminal fermentation and methane production of different seaweed species. Anim Feed Sci Technol 2017;228:1-12. [Google Scholar] | [Crossref] - Ramin M, Franco M, Roleda MY.
In vitro evaluation of utilisable crude protein and methane production for a diet in which grass silage was replaced by different levels and fractions of extracted seaweed proteins. Anim Feed Sci Technol 2019;255((July)):114225. [Google Scholar] | [Crossref] - Ramin M, Huhtanen P. Development of equations for predicting methane emissions from ruminants. J Dairy Sci 2013;96(4):2476-2493. [Google Scholar] | [Crossref]
- Özkan Gülzari, Lind V, Aasen IM, Steinshamn H. Effect of supplementing sheep diets with macroalgae species on
in vivo nutrient digestibility, rumen fermentation and blood amino acid profile. Animal 2019;13(12):2792-2801. [Google Scholar] | [Crossref] - Roleda MY, Marfaing H, Desnica N. Variations in polyphenol and heavy metal contents of wild-harvested and cultivated seaweed bulk biomass:health risk assessment and implication for food applications. Food Control 2019;95:121-134. [Google Scholar] | [Crossref]
- Jonsson NN, Ferguson HJ, McCartney CA. Postmortem observations on rumen wall histology and gene expression and ruminal and caecal content of beef cattle fattened on barley-based rations. Animal 2020;14(7):1447-1460. [Google Scholar] | [Crossref]
- Machado L, Magnusson M, Paul NA. Identification of bioactives from the red seaweed
Asparagopsis taxiformis that promote antimethanogenic activityin vitro . J Appl Phycol 2016;28(5):3117-3126. [Google Scholar] | [Crossref] - Kinley RD, Martinez-Fernandez G, Matthews MK. Mitigating the carbon footprint and improving productivity of ruminant livestock agriculture using a red seaweed. J Clean Prod 2020;259:120836. [Google Scholar] | [Crossref]
- Wahyuni DS, Jayanegara A, Wiryawan KG. Evaluation of several macroalgae species on methane emission and antioxidant activity based on
in vitro rumen fermentation characteristics. IOP Conf Ser Earth Environ Sci 2023. [Google Scholar] | [Crossref] - Puente-Rodríguez D, van Laar H, Veraart M. A circularity evaluation of new feed categories in The Netherlands—squaring the circle:a review. Sustainability 2022. [Google Scholar] | [Crossref]
- DeWeese KJ, Osborne MG. Understanding the metabolome and metagenome as extended phenotypes:the next frontier in macroalgae domestication and improvement. J World Aquac Soc 2021;52(5):1009-1030. [Google Scholar] | [Crossref]
- Kinley RD. The red macroalgae
Asparagopsis taxiformis is a potent natural antimethanogenic that reduces methane production duringin vitro fermentation with rumen fluid. Anim Prod Sci 2016;56:282-289. [Google Scholar] | [Crossref] - Molina-Alcaide E, Carro MD, Roleda MY.
In vitro ruminal fermentation and methane production of different seaweed species. Anim Feed Sci Technol 2017;228:1-12. [Google Scholar] | [Crossref] - Wanapat M, Viennasay B, Matra M. Supplementation of fruit peel pellet containing phytonutrients to manipulate rumen pH, fermentation efficiency, nutrient digestibility and microbial protein synthesis 2021. [Google Scholar] | [Crossref]
- Morais T, In A, Coutinho T, Ministro M. Seaweed potential in the animal feed:a review;3:1-24. [Google Scholar] | [Crossref]
- Coca FOCG, Gomes ENO, Junges L, Ítavo LCV, Nonato LM, Gomes FK, Ítavo CCBF, Difante GS, Dias AM. Protodioscin content, degradation kinetics, and
in vitro digestibility of Marandu palisadegrass hay as affected by cutting interval of the canopy. Trop Anim Sci J 2022;45(3):299-307. [Google Scholar] | [Crossref] - Morm S, Lunpha A, Pilajun R, Cherdthong A. Gas kinetics, rumen characteristics, and
in vitro degradability of varied levels of dried and fresh cassava leaf top fermented with cassava pulp. Trop Anim Sci J 2023;46(2):105-11. [Google Scholar] | [Crossref] - Guadayo GF, Rayos AA, Merca FE, Tandang AG, Loresco MM, Angeles AA. Prediction of
in situ ruminal degradability of forages in buffaloes using thein vitro gas production technique. Trop Anim Sci J 2019;42(2):128-36. [Google Scholar] | [Crossref]