


ABSTRACT
Background and Aim: The global expansion of antimicrobial resistance (AMR), particularly due to extended-spectrum β-lactamase (ESBL)-producing Escherichia coli, represents a critical threat to food safety and public health. Imported meat products are increasingly recognized as potential vehicles for the transboundary dissemination of multidrug-resistant (MDR) bacteria. However, data on the occurrence and molecular characteristics of ESBL-producing E. coli in imported frozen beef in Saudi Arabia remain limited. This study aimed to determine the prevalence, AMR profiles, and ESBL gene distribution among E. coli isolated from retail imported frozen beef.
Materials and Methods: A total of 78 imported frozen boneless beef samples were collected from retail shops and supermarkets in Dammam and Al Khobar, Eastern Province, Saudi Arabia. E. coli isolation was performed using Enterobacteriaceae enrichment broth followed by CHROMagar™ E. coli. Presumptive isolates were screened on CHROMagar™ ESBL and subjected to antimicrobial susceptibility testing against 21 antimicrobials using the disc diffusion method. Phenotypic confirmation of ESBL production was conducted using E-test ESBL strips. Molecular detection of ESBL-encoding genes (blaTEM, blaSHV, blaCTX-M-1, blaCTX-M-9, and blaCTX-M-15) was performed by polymerase chain reaction.
Results: From 78 beef samples, 390 E. coli isolates were recovered, of which 361 (92.5%) were presumptive ESBL producers on CHROMagar™ ESBL. Phenotypic confirmation showed that 319/361 (88.3%) isolates were ESBL-producing E. coli. Molecular analysis detected β-lactamase genes in 324/361 (89.7%) isolates, with blaCTX-M-1 and blaCTX-M-15 each identified in 87.5% of isolates. High resistance rates were observed to ampicillin (99.4%), cephalothin (97.2%), cephalexin (91.4%), ceftriaxone (81.9%), and cefotaxime (73.9%). MDR was detected in 97.2% of isolates.
Conclusion: Retail imported frozen beef in Saudi Arabia harbors an exceptionally high burden of MDR ESBL-producing E. coli, predominantly driven by CTX-M-1 and CTX-M-15 enzymes. These findings indicate that international beef supply chains may act as significant reservoirs for high-risk ESBL determinants, underscoring the need for strengthened surveillance, regulatory control, and One Health-based interventions to limit foodborne dissemination of AMR.
Keywords: antimicrobial resistance, beef safety, Escherichia coli, extended-spectrum beta-lactamase, foodborne pathogens, imported meat, multidrug resistance, One Health.
INTRODUCTION
Global health challenges have increasingly emerged as a consequence of rising antimicrobial resistance (AMR). Extended-spectrum β-lactamase-producing Escherichia coli (ESBL-producing E. coli) has been closely linked to this global threat [1]. E. coli has been isolated not only from clinical environments but also from food products imported from endemic regions. This bacterium is capable of hydrolyzing a broad range of β-lactam antibiotics, including penicillins and third-generation cephalosporins [2,3]. This hydrolytic activity disrupts the amide bonds within the four-membered β-lactam ring, thereby inactivating these antimicrobial agents [4].
β-lactamases are classified into four major groups based on molecular size and structural similarities within their active amino acid sites. Three groups belong to the serine β-lactamase classes A, C, and D, whereas the fourth group comprises metallo-β-lactamases (class B). ESBLs fall predominantly within class A and represent the most prevalent group [5]. These enzymes confer resistance to third-generation cephalosporins, such as cefotaxime, ceftriaxone, cefixime, and ceftazidime, which are critically important in both human and veterinary medicine. During the 1980s, a marked increase in ESBL prevalence was documented, largely associated with SHV and TEM variants that dominated early reports. However, following the first identification in Germany in 1989 of cefotaxime-resistant E. coli strains unrelated to TEM or SHV types, the cefotaxime-Munich (CTX-M) group emerged as the most widespread ESBL family worldwide and is now considered a major cause of infections in humans and animals within the Enterobacteriaceae [6, 7].
CTX-M enzymes originated from the mobilization of chromosomal β-lactamase genes in Kluyvera species, which subsequently became incorporated into mobile plasmids. Further diversification of CTX-M variants has occurred through point mutations driven by selective advantages under antibiotic pressure [8]. These enzymes act as natural cephalosporinases, exhibiting high activity against cefotaxime and ceftriaxone, and conferring resistance to other extended-spectrum cephalosporins, including cefepime and ceftazidime [9]. Although CTX-M genes are most commonly located on transferable conjugative plasmids flanked by insertion sequences that enhance mobility and expression, chromosomal integration has been reported with increasing frequency [10]. Based on amino acid sequence homology, CTX-M enzymes are classified into five phylogenetic families, each named after the first identified member [9], with more than 230 variants described to date [11].
The CTX-M family, particularly the CTX-M-1 and CTX-M-15 types, has become the dominant ESBL group globally, largely replacing earlier TEM and SHV types [12, 13]. These enzymes have been identified in both hospital-acquired and community-associated E. coli isolates [14]. ESBL-producing E. coli has also been recovered from various food products, including chicken, fish, and beef, often as a result of antibiotic use and misuse in animal production for disease control and growth promotion [15,16]. Imported meat products are increasingly recognized as important reservoirs and transmission routes for multidrug-resistant (MDR) bacteria, facilitating the silent spread of resistance genes into the human microbiota [17, 18]. The dissemination of these determinants is frequently mediated by mobile genetic elements, such as plasmids carrying resistance genes to multiple antimicrobial classes, posing a significant threat to public health [19].
Despite the growing global concern regarding AMR, data on ESBL-producing E. coli associated with imported meat products remain fragmented, particularly in the Middle East. Existing studies have largely focused on clinical isolates or locally produced foods, while systematic investigations addressing the prevalence, resistance profiles, and molecular characteristics of ESBL-producing E. coli in imported frozen beef are scarce. Moreover, limited information is available on the dominance and co-occurrence of key ESBL determinants, particularly CTX-M-1 and CTX-M-15, in retail beef products entering Saudi Arabia. The lack of integrated phenotypic and genotypic surveillance data hampers accurate risk assessment of MDR E. coli dissemination through international food supply chains and constrains the development of evidence-based food safety and AMR mitigation strategies within a One Health framework.
The present study aimed to investigate the occurrence of ESBL-producing E. coli in retail imported frozen beef marketed in the Eastern Province of Saudi Arabia. Specifically, this study sought to determine the prevalence of ESBL and MDR phenotypes among E. coli isolates, characterize their antimicrobial susceptibility patterns, and identify the distribution of major ESBL-encoding genes using molecular methods. By integrating phenotypic screening with genotypic confirmation, this work aims to provide baseline surveillance data on foodborne ESBL-producing E. coli and to clarify the potential role of imported frozen beef as a reservoir for high-risk AMR determinants.
MATERIALS AND METHODS
Ethical approval
This study was based exclusively on the examination of commercially available imported frozen beef samples, and no experiments involving live animals were conducted. Ethical review and approval were obtained from the Institutional Review Board, Imam Abdulrahman Bin Faisal University (IRB No. IRB 2014-04-189).
Study period and location
This study was conducted from April 2014 to April 2016 at Microbiology Research Laboratory, Department of Clinical Laboratory Sciences, Faculty of Applied Medical Sciences, Imam Abdulrahman Bin Faisal University. All E. coli isolates were stored in –80°C fridge in trypticase soy broth (TSB) containing 20% glycerol at Microbiology Research Laboratory. Between 2020 and 2021, all E. coli isolates stored in -800C fridge were analyzed using PCR to detect ESBL determinants genes.
Sample collection
A total of 78 imported frozen boneless beef samples were collected from retail shops and supermarkets located in Dammam and Al Khobar in the Eastern Province of Saudi Arabia. Samples were collected to investigate the presence of ESBL-producing E. coli and MDR among E. coli isolates. All samples were aseptically placed in sterile containers, maintained under ice, and transported to the microbiology research laboratories for microbiological analysis and genotypic characterization.
Isolation and identification of E. coli
E. coli was isolated from frozen beef samples using Enterobacteriaceae enrichment (EE) broth (CM013, Oxoid, Basingstoke, UK) and CHROMagar™ E. coli (CHROMagar, Paris, France). Briefly, 25 g of each beef sample was aseptically weighed and transferred into a sterile stomacher bag containing 225 mL of EE broth [20]. Samples were homogenized for 2 min using a laboratory blender (Seward Stomacher 400 Circulator, Seward, Worthing, UK) and incubated at 37°C for 24 h.
Following enrichment, 0.1 mL of the broth culture was streaked onto CHROMagar™ E. coli and incubated at 37°C for 24 h. Typical blue colonies suggestive of E. coli were selected and confirmed using API 20E identification strips (bioMérieux, Marcy-l’Étoile, France) along with standard oxidase and indole biochemical tests. Confirmed isolates were subsequently subcultured onto CHROMagar™ ESBL (CHROMagar, Paris, France). Colonies exhibiting dark pink to red coloration were presumptively identified as ESBL-producing E. coli and subjected to further antimicrobial susceptibility testing (AST). Phenotypic confirmation of ESBL production was performed using E-test ESBL strips, while genotypic confirmation was carried out by polymerase chain reaction (PCR).
AST
All E. coli isolates recovered on CHROMagar™ ESBL were tested for antimicrobial susceptibility using the disk diffusion method according to Clinical and Laboratory Standards Institute guidelines [21]. The antimicrobials tested included ciprofloxacin (CIP), cefotaxime (CTX), ceftazidime (CAZ), ceftriaxone (CRO), cephalexin (CFX), cefoxitin (FOX), cephalothin (KF), ampicillin (AM), amoxicillin/clavulanic acid (AUG), piperacillin (PIP), nalidixic acid (NA), norfloxacin (NOR), amikacin (AK), gentamicin (GM), kanamycin (K), tobramycin (TN), nitrofurantoin (FM), aztreonam (ATM), chloramphenicol (C), trimethoprim/sulfametho-xazole (SXT), and tetracycline (TE).
E. coli American Type Culture Collection 25922 was used as the reference strain for quality control. Results were interpreted according to CLSI criteria [21]. MDR was defined as non-susceptibility to at least one agent in three or more antimicrobial classes, following the criteria described by Magiorakos et al. [22].
Phenotypic screening for ESBL
Phenotypic confirmation of ESBL production was conducted using E-test ESBL strips (bioMérieux, Marcy-l’Étoile, France) after completion of AST. The E-test ESBL strips were used to determine minimum inhibitory concentrations (MICs) for ceftazidime/ceftazidime + clavulanic acid (TZ/TZL) and cefotaxime/ cefotaxime + clavulanic acid (CT/CTL). The strips generated concentration gradients for TZ (0.5–32 mg/L), TZ/TZL (0.064–4 mg/L plus 4 mg/L clavulanic acid), CT (0.25–16 mg/L), and CT/CTL (0.064–4 mg/L plus 4 mg/L clavulanic acid).
All selected E. coli isolates were tested according to the manufacturer’s instructions. ESBL production was confirmed when MIC values met the criteria: CT ≥ 0.5 mg/L with CT/CTL ≥ 8, and TZ ≥ 1 mg/L with TZ/TZL ≥ 8. ESBL activity was also inferred from the presence of “phantom” zones or deformation of inhibition ellipses. Results exceeding the measurable range of the strips were classified as non-determinable (ND).
DNA extraction and genotyping of β-lactamase genes
Genomic DNA was extracted using a boiling method as previously described by Yamani and Elhadi [23]. Briefly, 1.0 mL of an overnight E. coli culture grown in Luria–Bertani broth was centrifuged at 10,000 × g for 2 min. The pellet was resuspended in 1.0 mL of sterile distilled water and boiled at 100°C for 15 min, followed by centrifugation at 12,000 × g for 5 min. The supernatant containing DNA was transferred to a fresh tube and stored at −20°C until PCR analysis.
PCR was performed to detect blaCTX-M, blaCTX-M-1, blaCTX-M-15, blaCTX-M-9, blaTEM, and blaSHV genes using specific oligonucleotide primers as previously described [24–29]. Amplification reactions were carried out using GoTaq® Green Master Mix (Promega, Madison, WI, USA) in a T100 thermal cycler (Bio-Rad, Hercules, CA, USA). All primers were obtained from Eurofins Genomics (Ebersberg, Germany).
PCR products were separated on 1% agarose gels and visualized using a gel documentation system (G:BOX Chemi, Syngene, Cambridge, UK). Standard ESBL-producing E. coli and Klebsiella pneumoniae strains from previous studies were included as positive controls in each PCR run [20, 30].
RESULTS
Isolation of E. coli
The isolation outcomes for E. coli and ESBL-producing E. coli based on CHROMagar™ E. coli and CHROMagar™ ESBL are presented in Figure 1. Analysis of the 78 examined beef samples yielded 390 E. coli isolates showing typical blue colonies on CHROMagar™ E. coli (Figure 1). Of these, 361/390 (92.5%) isolates produced dark pink to red colonies after subculture on CHROMagar™ ESBL, indicating presumptive ESBL production (Figure 1).
Number of Escherichia coli isolates identified using CHROMagar™ E. coli, presumptive screening on CHROMagar™ extended-spectrum β-lactamase (ESBL), phenotypic confirmation of ESBL production using E-test ESBL strips, and genotypic confirmation by polymerase chain reaction.

Figure 1. Number of Escherichia coli isolates identified using CHROMagar™ E. coli, presumptive screening on CHROMagar™ extended-spectrum β-lactamase (ESBL), phenotypic confirmation of ESBL production using E-test ESBL strips, and genotypic confirmation by polymerase chain reaction.
Antimicrobial susceptibility profiles of isolates
The E. coli isolates recovered from imported frozen beef and tested against 21 antimicrobial agents exhibited variable susceptibility profiles (Table 1, Figure 2). The highest resistance rates among the 361 isolates were observed for AM (359/361, 99.4%), KF (351/361, 97.2%), CFX (330/361, 91.4%), PIP (315/361, 87.2%), CRO (296/361, 81.9%), and CTX (267/361, 73.9%).
Percentage of resistance, intermediate resistance, and susceptibility of Escherichia coli isolates (n = 361) tested against 21 antimicrobial drugs. CIP = Ciprofloxacin, CTX = Cefotaxime, CAZ = Ceftazidime, CRO = Ceftriaxone, CFX = Cephalexin, FOX = Cefoxitin, KF = Cephalothin, AM = Ampicillin, AUG = Augmentin, PIP = Piperacillin, NA = Nalidixic acid, NOR = Noroxin, AK = Amikacin, GM = Gentamicin, K = Kanamycin, TN = Tobramycin, FM = Nitrofurantoin, ATM = Aztreonam, C = Chloramphenicol, SXT = Trimethoprim/sulfamethoxazole, TE = Tetracycline.
In contrast, the lowest resistance rates were recorded for TN and FM (0.5% each), followed by AK and AUG (0.8% each) (Table 1). Notably, CAZ and FOX demonstrated comparatively low resistance levels of 3.0% and 1.6%, respectively. The highest proportion of intermediate resistance was unexpectedly detected for ATM (59.8%) (Table 1, Figure 2).
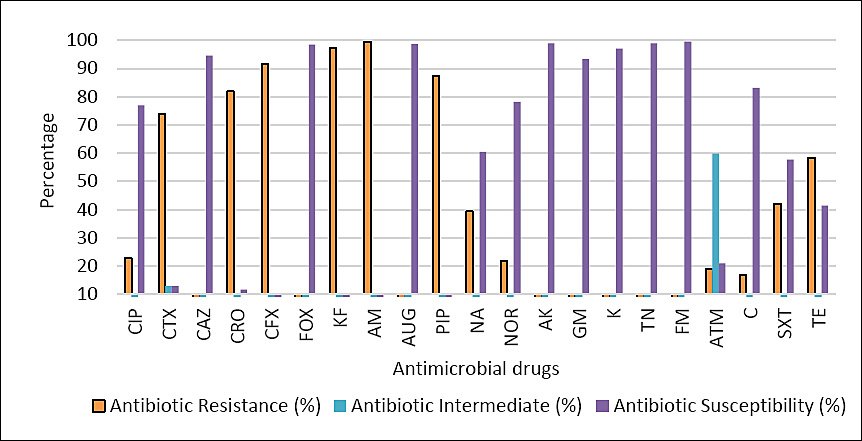
Figure 2. Percentage of resistance, intermediate resistance, and susceptibility of Escherichia coli isolates (n = 361) tested against 21 antimicrobial drugs. CIP = Ciprofloxacin, CTX = Cefotaxime, CAZ = Ceftazidime, CRO = Ceftriaxone, CFX = Cephalexin, FOX = Cefoxitin, KF = Cephalothin, AM = Ampicillin, AUG = Augmentin, PIP = Piperacillin, NA = Nalidixic acid, NOR = Noroxin, AK = Amikacin, GM = Gentamicin, K = Kanamycin, TN = Tobramycin, FM = Nitrofurantoin, ATM = Aztreonam, C = Chloramphenicol, SXT = Trimethoprim/sulfamethoxazole, TE = Tetracycline.
Table 1. Overall antibiotic susceptibility of Escherichia coli isolates (n = 361).
| Antibiotic class | Antibiotic | R (%) | I (%) | S (%) |
|---|---|---|---|---|
| Cephalosporins | Ciprofloxacin | 83 (22.9) | 0 | 278 (77.0) |
| Cefotaxime | 267 (73.9) | 47 (13.0) | 47 (13.0) | |
| Ceftazidime | 11 (3.0) | 9 (2.4) | 341 (94.4) | |
| Ceftriaxone | 296 (81.9) | 23 (6.3) | 42 (11.6) | |
| Cephalexin | 330 (91.4) | 10 (2.7) | 21 (5.8) | |
| Cefoxitin | 6 (1.6) | 0 | 355 (98.3) | |
| Cephalothin | 351 (97.2) | 7 (1.9) | 3 (0.8) | |
| Penicillins | Ampicillin | 359 (99.4) | 0 | 2 (0.8) |
| Amoxicillin/clavulanic acid | 3 (0.8) | 2 (0.5) | 356 (98.6) | |
| Piperacillin | 315 (87.2) | 14 (3.8) | 32 (8.8) | |
| Quinolones | Nalidixic acid | 142 (39.3) | 1 (0.2) | 218 (60.3) |
| Norfloxacin | 79 (21.8) | 0 | 282 (78.1) | |
| Aminoglycosides | Amikacin | 3 (0.8) | 1 (0.2) | 357 (98.8) |
| Gentamicin | 24 (6.6) | 0 | 337 (93.3) | |
| Kanamycin | 9 (2.4) | 2 (0.5) | 350 (96.9) | |
| Tobramycin | 2 (0.5) | 2 (0.5) | 357 (98.8) | |
| Nitrofuran | Nitrofurantoin | 2 (0.5) | 0 | 359 (99.4) |
| Monobactams | Aztreonam | 69 (19.1) | 216 (59.8) | 76 (21.0) |
| Amphenicol | Chloramphenicol | 61 (16.8) | 0 | 300 (83.1) |
| Sulfonamides | Trimethoprim/sulfamethoxazole | 152 (42.1) | 0 | 209 (57.8) |
| Tetracycline | Tetracycline | 211 (58.4) | 0 | 150 (41.5) |
R = Resistant, I = Intermediate, S = Susceptible.
MDR patterns
MDR, defined as non-susceptibility to at least one agent in three or more antimicrobial classes, was detected in 97.2% (351/361) of E. coli isolates. All resistance profiles involved non-susceptibility to more than one antimicrobial agent, except for a single isolate that remained susceptible to all tested drugs. The most extensive MDR profile included resistance to up to 16 antimicrobials (Supplementary Table).
The most frequently observed MDR pattern involved resistance to 6 antimicrobials, accounting for 18.2% (66/361) of isolates, followed by resistance to 12 antimicrobials, accounting for 8.8% (32/361) of isolates. Detailed MDR patterns are summarized in (Supplementary Table).
Prevalence of ESBL among E. coli isolates
Phenotypic confirmation using E-test ESBL strips demonstrated that 319/361 (88.3%) isolates recovered from CHROMagar™ ESBL exhibited an ESBL-positive phenotype (Figures 1 and 3). Molecular analysis detected β-lactamase-encoding genes in 324/361 (89.7%) isolates. The detected genes included blaTEM (n = 10, 2.7%), blaSHV (n = 89, 24.6%), blaCTX-M-1 group (n = 316, 87.5%), blaCTX-M-9 group (n = 17, 4.7%), and blaCTX-M-15 (n = 316, 87.5%), as illustrated in Figure 4.
Representative phenotypic patterns of growth inhibition in Escherichia coli isolates detected using the extended-spectrum β-lactamase (ESBL) E-test. (a) Distinct ESBL-positive result with a minimum inhibitory concentration (MIC) ratio of cefotaxime/cefotaxime + clavulanic acid (CT/CTL) > 32/0.047 = 32, accompanied by a rounded “phantom” inhibition zone beneath the ceftazidime (TZ) end, indicating ESBL production. (b) Rounded “phantom” inhibition zone beneath the CT end suggestive of ESBL activity, along with distortion of the TZ inhibition ellipse, also consistent with ESBL. (c) Distortion of the CT inhibition ellipse indicative of ESBL, together with a rounded “phantom” inhibition zone beneath the TZ end. (d) Deformation of both CT and TZ inhibition ellipses, characteristic of ESBL. (e) TZ inhibition ellipse deformation combined with a rounded “phantom” inhibition zone beneath the CT end, indicative of ESBL. (f) Deformation observed in both TZ and CT inhibition ellipses, confirming ESBL activity. (g and h) In cases where MIC values exceeded the measurable range of the test device, the results were classified as non-determinable.
Distribution and occurrence of β-lactamase-encoding genes among Escherichia coli isolates (n = 324).
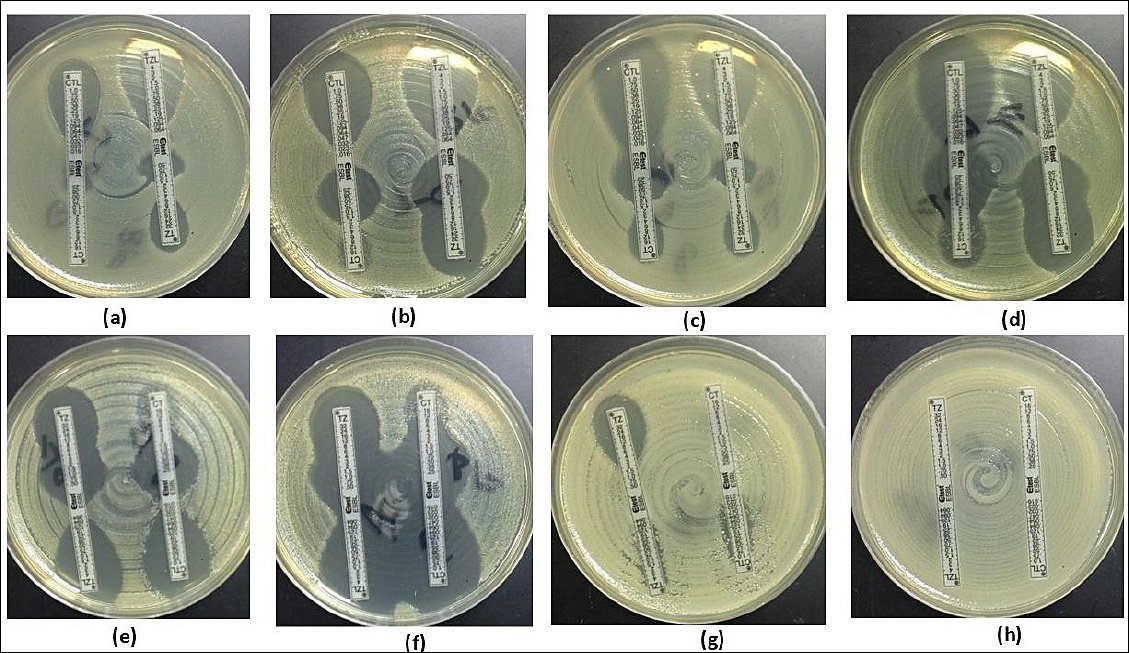
Figure 3. Representative phenotypic patterns of growth inhibition in Escherichia coli isolates detected using the extended-spectrum β-lactamase (ESBL) E-test. (a) Distinct ESBL-positive result with a minimum inhibitory concentration (MIC) ratio of cefotaxime/cefotaxime + clavulanic acid (CT/CTL) > 32/0.047 = 32, accompanied by a rounded “phantom” inhibition zone beneath the ceftazidime (TZ) end, indicating ESBL production. (b) Rounded “phantom” inhibition zone beneath the CT end suggestive of ESBL activity, along with distortion of the TZ inhibition ellipse, also consistent with ESBL. (c) Distortion of the CT inhibition ellipse indicative of ESBL, together with a rounded “phantom” inhibition zone beneath the TZ end. (d) Deformation of both CT and TZ inhibition ellipses, characteristic of ESBL. (e) TZ inhibition ellipse deformation combined with a rounded “phantom” inhibition zone beneath the CT end, indicative of ESBL. (f) Deformation observed in both TZ and CT inhibition ellipses, confirming ESBL activity. (g and h) In cases where MIC values exceeded the measurable range of the test device, the results were classified as non-determinable.
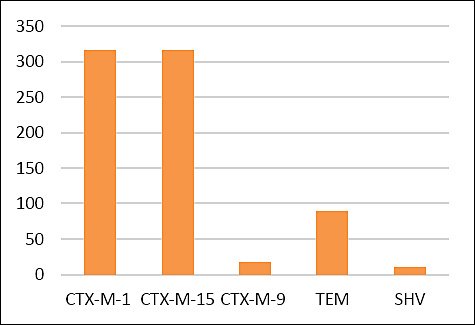
Figure 4. Distribution and occurrence of β-lactamase-encoding genes among Escherichia coli isolates (n = 324).
DISCUSSION
Livestock production and AMR
Livestock species play a vital role in the global food system and contribute substantially to economic, social, and cultural values. Meat remains a nutrient-dense food source worldwide, providing high-quality protein and essential micronutrients [31]. In addition to nutritional importance, livestock production represents a major economic sector in many regions [32]. However, intensive livestock farming faces significant challenges, foremost among them AMR, which poses a serious threat to both human and animal health. The excessive and inappropriate use of antimicrobials in livestock production has accelerated the emergence of resistant bacteria. These bacteria can be transmitted to humans through contaminated meat products, direct animal contact, or environmental pathways [33]. In many production systems, particularly in developing countries, antimicrobials are routinely administered to healthy animals at subtherapeutic doses for disease prevention and growth promotion, creating strong selective pressure for resistance development [34].
Role of E. coli as a foodborne reservoir
Among resistant bacteria, E. coli is one of the most prevalent species detected in animal-derived foods. E. coli commonly persists in livestock environments, originating from sources such as soil, contaminated feed, water runoff, or heavy metals [35]. Transmission to humans frequently occurs through consumption of cattle and poultry products. Once established in the human gut, resistance determinants are disseminated mainly through horizontal gene transfer. Unregulated antimicrobial use has contributed to the emergence of MDR E. coli, with resistance reported against multiple antimicrobial classes, including aminoglycosides, tetracyclines, β-lactams, fluoroquinolones, and third-generation cephalosporins [36–38]. Recent investigations of food-producing animals, including poultry, cattle, and farmed fish, demonstrated moderate to high prevalence of ESBL-producing E. coli, with the highest resistance rates consistently observed for TE and AM across geographic regions [38]. These studies also identified pandemic, high-risk zoonotic E. coli clones capable of contaminating humans and environmental niches.
ESBL-producing Enterobacteriaceae and CTX-M dominance
Members of the Enterobacteriaceae family include strains resistant to third-generation cephalosporins, particularly CTX-M group 1 variants, most notably blaCTX-M-1 and blaCTX-M-15, which are strongly associated with ESBL production in animals and humans and represent a growing global AMR challenge [38]. The high occurrence of ESBL-producing E. coli in imported frozen beef observed in this study highlights the food supply as an important source of MDR bacteria. Such contamination may facilitate transmission through human consumption and underscores the relevance of the One Health framework, linking AMR in livestock to food safety and healthcare systems. Increased prevalence of ESBL-producing Enterobacterales has been widely reported in food and clinical samples worldwide [39–42].
Molecular characteristics and global comparisons
In the present study, imported frozen beef showed a high prevalence of ESBL-producing E. coli, with 88.3% of isolates demonstrating phenotypic ESBL production and 89.7% harboring at least one β-lactamase gene. The most frequently detected genes were blaCTX-M-1 and blaCTX-M-15 (each 87.5%), while blaTEM, blaSHV, and blaCTX-M-9 groups were detected at lower frequencies [43–45]. These findings align with reports from Germany, where 93.4% of isolates from dairy and beef cattle carried CTX-M genes, predominantly blaCTX-M-1 [46]. Similarly, a comprehensive review from Taiwan identified E. coli as the most frequently isolated ESBL-producing species in animals, with blaTEM, blaSHV, and blaCTX-M as the dominant genes [47].
Public health relevance and One Health perspective
The predominance of CTX-M-1 and CTX-M-15 enzymes among isolates emphasizes the role of broad-spectrum antimicrobial misuse in driving AMR selection and spread [48]. The global expansion of CTX-M-producing E. coli has heightened concern regarding zoonotic transmission between animals and humans [39]. The World Health Organization has classified cephalosporin-resistant Enterobacteriaceae as critically important priority pathogens [49], and its Global Action Plan promotes a One Health approach to mitigate AMR [50]. Monitoring resistance markers is essential for identifying emerging trends and assessing health risks. In this study, isolates showed extensive resistance to β-lactams, particularly AM (99.4%), KF (97.2%), and CRO (81.9%), a resistance profile characteristic of ESBL-producing E. coli with frequent co-resistance to multiple antimicrobial classes [51]. The MDR rate observed here (97.2%) exceeds that reported in several international retail meat studies [17, 52, 53], underscoring the urgency for stricter antimicrobial regulation in livestock production.
Antimicrobial use in livestock and global trade implications
Antimicrobials are widely used in livestock farming for disease prevention, treatment, and growth promotion [54]. Their overuse results in environmental residues and sustained selective pressure that favors resistant strains through successive mutation cycles [55]. Approximately half of global antimicrobial production is consumed in livestock, with the United States, China, Australia, and Brazil accounting for 58% of use in 2020 [56]. Antimicrobial consumption is highest in Asia, followed by Oceania and parts of Europe, with TE being the most extensively used agent globally [57]. Such intensive use contributes to the emergence of ESBL-producing bacteria in animal-derived foods [57, 58].
International trade and dissemination of resistance
The Food and Agriculture Organization has highlighted AMR as a global priority in its action plans [59], noting that countries lacking robust regulatory frameworks are at highest risk. Globalization of the food trade has accelerated cross-border dissemination of AMR, as foods carrying resistance genes on mobile elements are imported into countries with strict domestic regulations [60]. Livestock production in regions with limited oversight, combined with improper waste management, further amplifies environmental dissemination [61]. Similar concerns apply to aquaculture systems operating under weak regulatory conditions [62, 63]. Assessing AMR risks in livestock remains challenging due to limited data on antimicrobial use and the complexity of production systems [62, 63].
Implications of resistance patterns in imported beef
In this study, resistance among E. coli isolates was observed against cephalosporins, penicillins, quinolones, and sulfonamides, with notably high intermediate resistance to ATM (59.8%). Although ATM is primarily reserved for clinical use, reduced susceptibility in foodborne isolates may indicate horizontal gene transfer or co-selection driven by other antimicrobials [64, 65]. Such mechanisms may facilitate the introduction of resistant pathogens into importing countries, including Saudi Arabia. Despite growing evidence of AMR in imported frozen beef and fishery products, data from Saudi Arabia remain limited [20, 36, 64–66]. Imported frozen beef and other animal-derived foods may therefore act as reservoirs for clinically relevant resistance determinants, including ESBLs, plasmid-mediated quinolone resistance, colistin resistance, and carbapenem resistance, posing ongoing risks to the food chain and public health [67].
CONCLUSION
This study demonstrated an exceptionally high burden of ESBL-producing E. coli in retail imported frozen beef, with 88.3% of isolates confirmed phenotypically as ESBL producers and 89.7% harboring at least one β-lactamase gene. The ESBL profile was dominated by blaCTX-M-1 and blaCTX-M-15, each detected in 87.5% of isolates, while blaTEM, blaSHV, and blaCTX-M-9 groups were identified at lower frequencies. AST revealed extensive resistance to β-lactams, particularly AM, KF, and CRO, and an alarmingly high MDR rate of 97.2%, with some isolates resistant to up to 16 antimicrobial agents.
The detection of widespread MDR ESBL-producing E. coli in imported frozen beef highlights a substantial food safety concern and indicates that international meat supply chains may act as vehicles for the transboundary dissemination of high-risk AMR determinants. The predominance of CTX-M-1 and CTX-M-15 underscores the potential for zoonotic transfer of clinically relevant resistance genes through the food chain. These findings emphasize the need for strengthened import screening, harmonized AMR surveillance, and stricter regulation of antimicrobial use in food-producing animals within a One Health framework.
Key strengths of this study include the integrated use of culture-based isolation, phenotypic ESBL confirmation, comprehensive AST, and molecular genotyping, providing robust and complementary evidence of ESBL and MDR occurrence. The large number of isolates recovered per sample enhanced detection sensitivity, and the focus on imported frozen beef addresses a critical but underexplored AMR pathway in the region.
This study was limited to retail imported frozen beef from a single province, which may not fully reflect national or seasonal variability. Whole-genome sequencing and plasmid characterization were not performed, limiting insights into clonal relatedness, mobile genetic elements, and transmission dynamics. Additionally, quantitative exposure assessment and direct linkage to human infections were beyond the scope of this work.
Future studies should expand surveillance to multiple regions and food matrices, integrate whole-genome sequencing to elucidate transmission pathways, and assess the contribution of specific exporting countries and production systems. Risk assessment studies linking foodborne ESBL-producing E. coli to human colonization and infection are also warranted to inform targeted interventions.
Overall, this study provides compelling evidence that imported frozen beef constitutes a significant reservoir of MDR ESBL-producing E. coli, predominantly driven by CTX-M-1 and CTX-M-15 enzymes. These findings reinforce the urgency for coordinated One Health strategies, enhanced regulatory oversight, and sustained AMR monitoring to mitigate the public health risks associated with global food trade.
DATA AVAILABILITY
The supplementary data can be made available from the corresponding author upon request.
AUTHORS’ CONTRIBUTIONS
LZY: Methodology, data analysis, and drafted and revised the manuscript. NE: Conceptualization, methodology, sample collection, data analysis, and drafted and revised the manuscript. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
This research was funded by the Deanship of Scientific Research, Imam Abdulrahman Bin Faisal University (grant number 2014191).
REFERENCES
- Woerther P-L, Burdet C, Chachaty E, Andremont A. Trends in human fecal carriage of extended-spectrum β-lactamases in the community:Toward the globalization of CTX-M. Clin Microbiol Rev 2013;26(4):744-758. [Google Scholar] | [Crossref]
- Madec J-Y, Haenni M, Nordmann P, Poirel L. Extended-spectrum β-lactamase/AmpC- and carbapenemase-producing Enterobacteriaceae in animals:A threat for humans?. Clin Microbiol Infect 2017;23(11):826-833. [Google Scholar] | [Crossref]
- Ribeiro LF, Nespolo NM, Rossi GAM, Fairbrother JM. Exploring extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli in food-producing animals and animal-derived foods. Pathogens 2024;13(4):346. [Google Scholar] | [Crossref]
- Bush K, Bradford PA. β-Lactams and β-lactamase inhibitors:An overview. Cold Spring Harb Perspect Med 2016;6((8)):a025247. [Google Scholar] | [Crossref]
- Bush K, Jacoby GA. Updated functional classification of β-lactamases. Antimicrob Agents Chemother 2010;54(3):969-976. [Google Scholar] | [Crossref]
- Bush K. Classification of beta-lactamases:Groups 1, 2a, 2b, and 2b′. Antimicrob Agents Chemother 1989;33(3):264-270. [Google Scholar] | [Crossref]
- Castanheira M, Simner PJ, Bradford PA. Extended-spectrum β-lactamases:An update on their characteristics, epidemiology and detection. JAC Antimicrob Resist 2021;3(3):dlab092. [Google Scholar] | [Crossref]
- Cantón R, González-Alba JM, Galán JC. CTX-M enzymes:Origin and diffusion. Front Microbiol 2012;3:110. [Google Scholar] | [Crossref]
- Bonnet R. Growing group of extended-spectrum β-lactamases:The CTX-M enzymes. Antimicrob Agents Chemother 2004;48(1):1-14. [Google Scholar] | [Crossref]
- Hussain HI, Aqib AI, Seleem MN, Shabbir MA, Hao H, Iqbal Z. Genetic basis of molecular mechanisms in β-lactam-resistant gram-negative bacteria. Microb Pathog 2021;158:105040. [Google Scholar] | [Crossref]
- Mendonça J, Guedes C, Silva C, Sá S, Oliveira M, Accioly G. New CTX-M group conferring β-lactam resistance:A compendium of phylogenetic insights from biochemical, molecular, and structural biology. Biology (Basel) 2022;11(2):256. [Google Scholar] | [Crossref]
- Bevan ER, Jones AM, Hawkey PM. Global epidemiology of CTX-M β-lactamases:Temporal and geographical shifts in genotype. J Antimicrob Chemother 2017;72(8):2145-2155. [Google Scholar] | [Crossref]
- Rozwandowicz M, Brouwer MSM, Fischer J, Wagenaar JA, Gonzalez-Zorn B, Guerra B. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J Antimicrob Chemother 2018;73(5):1121-1137. [Google Scholar] | [Crossref]
- Cantón R, Akóva M, Carmeli Y, Giske CG, Glupczynski Y, Gniadkowski M. Rapid evolution and spread of carbapenemases among Enterobacteriaceae in Europe. Clin Microbiol Infect 2012;18(5):413-431. [Google Scholar] | [Crossref]
- Tang KL, Caffrey NP, Nóbrega DB, Cork SC, Ronksley PE, Barkema HW. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings:A systematic review and meta-analysis. Lancet Planet Heal 2017;1(8):e316. [Google Scholar] | [Crossref]
- Michael CA, Dominey-Howes D, Labbate M. The antimicrobial resistance crisis:Causes, consequences, and management. Front Public Heal 2014;2:145. [Google Scholar] | [Crossref]
- Zurfluh K, Nüesch-Inderbinen M, Morach M, Zihler Berner A, Hächler H, Stephan R. Extended-spectrum-β-lactamase-producing Enterobacteriaceae isolated from vegetables imported from the Dominican Republic, India, Thailand, and Vietnam. Appl Environ Microbiol 2015;81(9):3115-3120. [Google Scholar] | [Crossref]
- Endale H, Mathewos M, Abdeta D. Potential causes of spread of antimicrobial resistance and preventive measures in One Health perspective:A review. Infect Drug Resist 2023;16:7515-7545. [Google Scholar] | [Crossref]
- Partridge SR, Kwong SM, Firth N, Jensen SO. Mobile genetic elements associated with antimicrobial resistance. Clin Microbiol Rev 2018;31(4):e00088-17. [Google Scholar] | [Crossref]
- Elhadi N. Prevalence of extended-spectrum-β-lactamase-producing Escherichia coli in imported frozen freshwater fish in Eastern Province of Saudi Arabia. Saudi J Med Med Sci 2016;4(1):19-25. [Google Scholar] | [Crossref]
- Performance standards for antimicrobial susceptibility testing. Wayne, PA: Clinical and Laboratory Standards Institute; 2017. [Google Scholar]
- Magiorakos A-P, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria:An international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect 2012;18(3):268-281. [Google Scholar] | [Crossref]
- Yamani LZ, Elhadi N. Virulence characteristics, antibiotic resistance patterns and molecular typing of enteropathogenic Escherichia coli (EPEC) isolates in Eastern Province of Saudi Arabia, 2013–2014. Infect Drug Resist 2022;15:6763-6772. [Google Scholar] | [Crossref]
- Saladin M, Cao VTB, Lambert T, Donay J-L, Herrmann J-L, Ould-Hocine Z. Diversity of CTX-M β-lactamases and their promoter regions from Enterobacteriaceae isolated in three Parisian hospitals. FEMS Microbiol Lett 2002;209(2):161-168. [Google Scholar] | [Crossref]
- Jeong SH, Bae IK, Kwon SB, Lee JH, Song JS, Jung HI. Dissemination of transferable CTX-M-type extended-spectrum β-lactamase-producing Escherichia coli in Korea. J Appl Microbiol 2005;98(4):921-927. [Google Scholar] | [Crossref]
- Muzaheed, Doi Y, Adams-Haduch JM, Endimiani A, Sidjabat HE, Gaddad SM. High prevalence of CTX-M-15-producing Klebsiella pneumoniae among inpatients and outpatients with urinary tract infection in Southern India. J Antimicrob Chemother 2008;61(6):1393-1394. [Google Scholar] | [Crossref]
- Costa D, Poeta P, Sáenz Y, Vinué L, Rojo-Bezares B, Jouini A. Detection of Escherichia coli harbouring extended-spectrum β-lactamases of the CTX-M, TEM and SHV classes in faecal samples of wild animals in Portugal. J Antimicrob Chemother 2006;58(6):1311-1312. [Google Scholar] | [Crossref]
- Arpin C, Dubois V, Coulange L, André C, Fischer I, Noury P. Extended-spectrum β-lactamase-producing Enterobacteriaceae in community and private health care centers. Antimicrob Agents Chemother 2003;47(11):3506-3514. [Google Scholar] | [Crossref]
- De Gheldre Y, Avesani V, Berhin C, Delmée M, Glupczynski Y. Evaluation of Oxoid combination discs for detection of extended-spectrum β-lactamases. J Antimicrob Chemother 2003;52(4):591-597. [Google Scholar] | [Crossref]
- Abdalhamid B, Elhadi N, Albunayan S, Alsamman K, Aljindan R. First description of methyltransferases in extensively drug-resistant Klebsiella pneumoniae isolates from Saudi Arabia. J Med Microbiol 2017;66(7):859-863. [Google Scholar] | [Crossref]
- Ederer P, Baltenweck I, Blignaut JN, Moretti C, Tarawali S. Affordability of meat for global consumers and the need to sustain investment capacity for livestock farmers. Anim Front 2023;13(2):45-60. [Google Scholar] | [Crossref]
- Enshaie E, Nigam S, Patel S, Rai V. Livestock antibiotics use and antimicrobial resistance. Antibiotics 2025;14(6):621. [Google Scholar] | [Crossref]
- Almansour AM, Alhadlaq MA, Alzahrani KO, Mukhtar LE, Alharbi AL, Alajel SM. The silent threat:Antimicrobial-resistant pathogens in food-producing animals and their impact on public health. Microorganisms 2023;11(9):2127. [Google Scholar] | [Crossref]
- van den Honert MS, Gouws PA, Hoffman LC. Escherichia coli antibiotic resistance patterns from co-grazing and non-co-grazing livestock and wildlife species from two farms in the Western Cape, South Africa. Antibiotics 2021;10(6):618. [Google Scholar] | [Crossref]
- Xu C, Kong L, Liao Y, Tian Y, Wu Q, Liu H. Mini-review:Antibiotic-resistant Escherichia coli from farm animal-associated sources. Antibiotics 2022;11(11):1535. [Google Scholar] | [Crossref]
- Alhabib I, Elhadi N. Antimicrobial resistance pattern of Escherichia coli isolated from imported frozen shrimp in Saudi Arabia. PeerJ 2024;12:e18689. [Google Scholar] | [Crossref]
- Silva A, Silva V, Pereira JE, Maltez L, Igrejas G, Valentão P. Antimicrobial resistance and clonal lineages of Escherichia coli from food-producing animals. Antibiotics 2023;12(6):1061. [Google Scholar] | [Crossref]
- Higgins O, Chueiri A, O'Connor L, Lahiff S, Burke L, Morris D. Portable differential detection of CTX-M ESBL gene variants blaCTX-M-1 and blaCTX-M-15 from Escherichia coli isolates and animal fecal samples. Microbiol Spectr 2023;11(1):e03316-22. [Google Scholar] | [Crossref]
- Dokuta S, Zhang X, Jeeno P, Hongjaisee S, Yadoung S, Khamnoi P. ESBL-producing Enterobacterales in food and clinical samples:Antimicrobial resistance organisms and genes in Chiang Mai, Thailand. Sci Rep 2025;15(1):23886. [Google Scholar] | [Crossref]
- Mangroliya DB, Adhyaru HJ, Kabariya JH, Ramani VM. High-risk AMR genes blaCMY-42, blaCTX-M-15, and blaNDM-5 in raw milk:A foodborne transmission concern. The Microbe 2025;100377. [Google Scholar] | [Crossref]
- Damianos A, Tsitsos A, Economou V, Gioula G, Haidich A-B. Systematic review and meta-analysis of the occurrence of ESBL-producing Escherichia coli and Salmonella spp in foods of animal origin in Europe. Food Control 2025;171:111127. [Google Scholar] | [Crossref]
- Husna A, Rahman MM, Badruzzaman ATM, Sikder MH, Islam MR, Rahman MT. Extended-spectrum β-lactamases (ESBL):Challenges and opportunities. Biomedicines 2023;11((11)):2937. [Google Scholar] | [Crossref]
- Kelbert L, Barmettler K, Biggel M, Stephan R, Nüesch-Inderbinen M. Occurrence and characteristics of extended-spectrum β-lactamase-producing Escherichia coli in Swiss and imported retail chicken meat. J Glob Antimicrob Resist 2025. [Google Scholar] | [Crossref]
- Mandujano-Hernández A, Martínez-Vázquez AV, Paz-González AD, Herrera-Mayorga V, Sánchez-Sánchez M, Lara-Ramírez EE. The global rise of ESBL-producing Escherichia coli in the livestock sector:A five-year overview. Animals 2024;14(17):2490. [Google Scholar] | [Crossref]
- Schmid A, Hörmansdorfer S, Messelhäusser U, Käsbohrer A, Sauter-Louis C, Mansfeld R. Prevalence of extended-spectrum β-lactamase-producing Escherichia coli on Bavarian dairy and beef cattle farms. Appl Environ Microbiol 2013;79(9):3027-3032. [Google Scholar] | [Crossref]
- Tseng C-H, Liu C-W, Liu P-Y. Extended-spectrum β-lactamases (ESBL)-producing bacteria in animals. Antibiotics 2023;12(4):661. [Google Scholar] | [Crossref]
- Ye Z, Li M, Jing Y, Liu K, Wu Y, Peng Z. What are the drivers triggering antimicrobial resistance emergence and spread?Outlook from a One Health perspective. Antibiotics 2025;14(6):543. [Google Scholar] | [Crossref]
- Sati H, Carrara E, Savoldi A, Hansen P, Garlasco J, Campagnaro E. The WHO bacterial priority pathogens list 2024:A prioritisation study to guide research, development, and public health strategies against antimicrobial resistance. Lancet Infect Dis 2025. [Google Scholar] | [Crossref]
- Action against antimicrobial resistance requires a One Health approach. Geneva: World Health Organization; 2024. [Google Scholar]
- Rus A, Bucur I-M, Imre K, Tirziu AT, Ivan AA, Gros RV. Phenotypic and genotypic characterization of ESBL and AmpC β-lactamase-producing Escherichia coli isolates from poultry in Northwestern Romania. Antibiotics 2025;14(6):578. [Google Scholar] | [Crossref]
- Mgaya FX, Matee MI, Muhairwa AP, Hoza AS. Occurrence of multidrug-resistant Escherichia coli in raw meat and cloaca swabs in poultry processed in slaughter slabs in Dar es Salaam, Tanzania. Antibiotics 2021;10(4):343. [Google Scholar] | [Crossref]
- Ren X, Yang D, Yang Z, Li Y, Yang S, Li W. Prevalence and antimicrobial susceptibility of foodborne pathogens from raw livestock meat in China, 2021. Microorganisms 2024;12(11):2157. [Google Scholar] | [Crossref]
- Odey TOJ, Tanimowo WO, Afolabi KO, Jahid IK, Reuben RC. Antimicrobial use and resistance in food animal production:Food safety and associated concerns in sub-Saharan Africa. Int Microbiol 2024;27(1):1-23. [Google Scholar] | [Crossref]
- Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP. Global trends in antimicrobial use in food animals. Proc Natl Acad Sci USA 2015;112(18):5649-5654. [Google Scholar] | [Crossref]
- Mulchandani R, Wang Y, Gilbert M, Van Boeckel TP. Global trends in antimicrobial use in food-producing animals:2020 to 2030. PLOS Glob Public Heal 2023;3(2):e0001305. [Google Scholar] | [Crossref]
- Kürekci C, Yüksel M, Celil Ozaslan BG, Tan S, Jäckel C, Grobbel M. Diversity and resistance profiles of ESBL-producing gram-negative bacteria from dairy farms in Southern Türkiye. Antibiotics 2024;13(12):1134. [Google Scholar] | [Crossref]
- The FAO action plan on antimicrobial resistance 2016–2020. Rome: FAO; 2016. [Google Scholar]
- Musuka G, Machakwa J, Mano O, Iradukunda PG, Gashema P, Moyo E. Antimicrobial resistance and its impact on food safety determinants along the beef value chain in sub-Saharan Africa:A scoping review. Trop Med Infect Dis 2025;10(3):82. [Google Scholar] | [Crossref]
- Khmaissa M, Zouari-Mechichi H, Sciara G, Record E, Mechichi T. Pollution from livestock farming antibiotics:An emerging environmental and human health concern. J Hazard Mater Adv 2024;13:100410. [Google Scholar] | [Crossref]
- Gilbert W, Thomas LF, Coyne L, Rushton J. Mitigating the risks posed by intensification in livestock production:The examples of antimicrobial resistance and zoonoses. Anim 2021;15(2):100123. [Google Scholar] | [Crossref]
- Salam MA, Al-Amin MY, Salam MT, Pawar JS, Akhter N, Rabaan AA. Antimicrobial resistance:A growing serious threat for global public health. Healthcare 2023;11:1946. [Google Scholar] | [Crossref]
- Kiskó G, Bajramović B, Elzhraa F, Erdei-Tombor P, Dobó V, Mohácsi-Farkas C. The invisible threat of antibiotic resistance in food. Antibiotics 2025;14(3):250. [Google Scholar] | [Crossref]
- Adzitey F, Huda N, Shariff AHM. Phenotypic antimicrobial susceptibility of Escherichia coli from raw meats, ready-to-eat meats, and their related samples in One Health context. Microorganisms 2021;9(2):326. [Google Scholar] | [Crossref]
- Almulhim A, Alomar A, Alhabib I, Yamani LZ, Elhadi N. Isolation of Shiga toxin-producing Escherichia coli O157 and non-O157 from retail imported frozen beef marketed in Saudi Arabia using immunomagnetic separation and multiplex PCR. Germs 2024;14(4):352. [Google Scholar] | [Crossref]
- Alsultan A, Elhadi N. Evaluation of ERIC-PCR method for determining genetic diversity among Escherichia coli isolated from human and retail imported frozen shrimp and beef. Int J Food Contam 2022;9(1):12. [Google Scholar] | [Crossref]
- Elhadi N. Prevalence and antimicrobial resistance of Salmonella spp. in raw retail frozen imported freshwater fish to Eastern Province of Saudi Arabia. Asian Pac J Trop Biomed 2014;4((3)):234-238. [Google Scholar] | [Crossref]
- Jung D, Morrison BJ, Rubin JE. A review of antimicrobial resistance in imported foods. Can J Microbiol 2022;68(1):1-15. [Google Scholar] | [Crossref]