


ABSTRACT
Background and Aim: Antimicrobial resistance (AMR) in foodborne bacteria presents a significant threat to public health, especially in countries with intensive livestock production systems. Pig farming is a major source of animal protein in Thailand and is recognized as an important reservoir of antimicrobial-resistant bacteria. Escherichia coli is commonly used as an indicator organism for monitoring AMR, including extended-spectrum β-lactamase (ESBL) production and pathogenic potential. This study aimed to assess the frequency of AMR, multidrug-resistant (MDR), ESBL determinants, and virulence genes in E. coli isolates collected from slaughterhouses and fresh markets in central Thailand.
Materials and Methods: A total of 498 archived E. coli isolates were analyzed, including 236 isolates from slaughterhouses (feces and carcasses) and 262 isolates from fresh markets (pork and cutting boards). Antimicrobial susceptibility testing was performed against 18 antimicrobial agents using the disk diffusion method. MDR was defined as resistance to three or more antimicrobial classes. ESBL production was identified through phenotypic confirmatory tests, and ESBL-producing isolates were screened for blaTEM, blaCTX-M, and blaSHV genes by multiplex polymerase chain reaction. All isolates were further examined for select virulence genes linked to major E. coli pathotypes.
Results: Overall, 97.4% of E. coli isolates showed resistance to at least one antimicrobial agent, and 87.3% were classified as MDR. ESBL-producing E. coli made up 23.5% of all isolates, with a significantly higher prevalence in slaughterhouses compared to fresh markets (p < 0.05). Among ESBL producers, 97.4% exhibited MDR phenotypes. Most (89.7%) of the ESBL-producing isolates carried at least one bla gene, with blaTEM being the most common, followed by blaCTX-M. Virulence genes were detected at a low frequency (3.2%), mainly involving eaeA, lt, and stp.
Conclusion: The high prevalence of AMR, MDR, and ESBL-producing E. coli throughout the pork production chain highlights slaughterhouses and fresh markets as key points for the spread of resistant bacteria. These findings emphasize the need for stronger antimicrobial stewardship, better hygiene practices, and ongoing AMR surveillance within the One Health approach to reduce public health risks linked to pork consumption.
Keywords: antimicrobial resistance, extended-spectrum beta-lactamase, Escherichia coli, food safety, multidrug resistance, pork production chain, slaughterhouse contamination, Thailand, virulence genes.
INTRODUCTION
Antimicrobial resistance (AMR) is widely acknowledged as a major global public health threat, causing approximately 700,000 deaths worldwide each year [1]. The widespread use of antimicrobial agents in both human healthcare and animal production systems is a key factor in the emergence and spread of AMR. In Thailand, pigs are one of the main food-producing animals and have been identified as a significant reservoir of antimicrobial-resistant bacteria [2, 3]. In pig production systems, antimicrobials are commonly used for both therapeutic and growth-promotion purposes. Previous studies have reported that about 78%–99% of Escherichia coli isolates from pigs show AMR phenotypes [4]. Antimicrobial-resistant bacteria colonizing food animals can be transmitted to humans through the consumption and handling of contaminated food products, direct contact with animals, and environmental spread of resistant organisms [2].
Additionally, a rising trend in the emergence of multidrug-resistant (MDR) bacteria in food animals has been observed [5]. Numerous studies have emphasized the increasing prevalence of extended-spectrum β-lactamase (ESBL)-producing E. coli, which represents a global public health crisis. ESBLs are enzymes that provide resistance to extended-spectrum cephalosporins, such as cefotaxime, ceftazidime, and ceftriaxone, as well as to monobactams [6]. These enzymes have been increasingly detected among members of the Enterobacteriaceae family, especially E. coli strains isolated from food-producing animals [4]. The most commonly identified ESBL-associated genes include blaCTX and blaTEM. Additionally, various ESBL genes have been found in E. coli from diverse animal-derived food sources [7]. These reservoirs are particularly concerning because they may harbor E. coli strains resistant to critically important antimicrobials and help facilitate their transmission to humans via the food chain.
E. coli is also the etiological agent of colibacillosis, a common infectious disease affecting both pigs and humans [8]. In pigs, colibacillosis poses a significant challenge to the industry due to its association with increased morbidity and mortality [9]. Enterotoxigenic E. coli (ETEC), enteropathogenic E. coli (EPEC), and Shiga toxin–producing E. coli (STEC) are the main causes of neonatal diarrhea, post-weaning diarrhea, and edema disease in pigs [10]. These pathotypes are not only economically important in pig production but also pose significant foodborne public health risks, having been linked to severe and sometimes deadly outbreaks worldwide. As a result, pigs are widely recognized as important reservoirs of pathogenic E. coli, with the potential to contaminate pork products and transmit infections to consumers [8].
In Thailand, pig production mainly occurs in the central provinces around Bangkok, which together make up about 36%–40% of the country’s total pig output [11].
Despite growing recognition of AMR and ESBL-producing E. coli as major threats to food safety and public health, significant knowledge gaps remain along the pork production chain in Thailand. Most existing studies have focused on farm-level surveillance or retail meat products, with limited integration of multiple critical points within the same production process. In particular, systematic data linking slaughterhouse environments (fecal and carcass contamination) with downstream fresh-market settings (pork and food-contact surfaces such as cutting boards) are scarce. This lack of integrated surveillance hampers the ability to identify key contamination points and understand how antimicrobial-resistant E. coli persist and spread from primary processing to retail environments.
Furthermore, while several studies have reported the prevalence of AMR or ESBL-producing E. coli in food animals, fewer investigations have simultaneously characterized MDR patterns, ESBL determinants, and virulence gene profiles within the same isolate collection. Such combined analyses are crucial to distinguish between commensal reservoirs of resistance and strains with potential pathogenicity. In addition, region-specific data from central Thailand, where pig production and pork distribution are highly concentrated, remain limited, especially concerning the comparative burden of resistance and ESBL genes between slaughterhouses and fresh markets. The lack of this information limits evidence-based risk assessment and the development of targeted intervention strategies across the pork production chain.
This study aimed to thoroughly examine AMR, MDR, ESBL production, and virulence gene profiles of E. coli strains collected from key points along the pork production chain in central Thailand. Specifically, it sought to (i) identify AMR patterns and the prevalence of MDR in E. coli isolates from slaughterhouses and fresh markets; (ii) determine the frequency of ESBL-producing E. coli and analyze the distribution of major ESBL-related genes (blaTEM, blaCTX-M, and blaSHV); (iii) investigate the presence of specific virulence genes linked to clinically and epidemiologically significant E. coli pathotypes; and (iv) compare resistance profiles and gene distribution between the upstream (slaughterhouse) and downstream (fresh-market) stages of the pork supply chain. By achieving these objectives, the study aims to provide baseline data to support risk-reduction strategies, antimicrobial stewardship, and enhanced food-safety measures within a One Health approach.
MATERIALS AND METHODS
Ethical approval
This study did not involve live animals or human participants. All E. coli isolates analyzed were obtained from a preserved stock culture collection maintained by the Faculty of Public Health at Thammasat University, Thailand, which were originally collected during routine microbiological surveillance activities conducted in 2017–2018. No animals were handled, restrained, sampled, or subjected to any intervention by the research team. According to Thai national regulations, studies using archived bacterial isolates do not require approval from an Institutional Animal Care and Use Committee or a Human Research Ethics Committee. Institutional exemption was granted by the Ethics Review Committee of the Faculty of Public Health at Thammasat University, confirming that the study was exempt from ethical review because it involved only preserved microorganisms and no identifiable animal, human, or personal data (Exemption ID: 0516.71/706). All laboratory procedures and data handling followed institutional biosafety guidelines, Thailand Ministry of Public Health regulations, and international standards for research involving microbial isolates.
Study design, period, and location
This study used a retrospective laboratory-based design with E. coli isolates collected along the pork production chain, including three slaughterhouses and four fresh markets in a central province of Thailand. Sample collection took place during 2017–2018. All E. coli isolates were stored at −80°C in glycerol stock cultures at the Faculty of Public Health, Thammasat University. Between 2019 and 2020, representative isolates from the stored stocks were selected for laboratory analysis to assess AMR profiles, ESBL determinants, and virulence genes.
Origin and selection of E. coli isolates
E. coli isolates were collected from slaughterhouses supervised by the Department of Livestock Development, Ministry of Agriculture and Cooperatives, and from large fresh markets in a central province of Thailand. All samples were initially identified as E. coli using standard bacterial culture techniques and biochemical tests, then stored at –80°C in glycerol. A total of 509 E. coli isolates were recovered from the preserved cultures. Of these, 498 isolates were successfully revived and included in the study, while 11 isolates could not be cultured. The analyzed isolates included samples from slaughterhouses (117 fecal and 119 carcass isolates) and from fresh markets (116 pork and 146 cutting board isolates).
Bacterial identification
All 498 E. coli isolates were recovered from preserved stock cultures and verified for purity by streaking onto MacConkey agar (Merck, Darmstadt, Germany), followed by incubation at 37°C for 18–24 h. Typical E. coli colonies appeared as pink colonies with a surrounding precipitation zone. Presumptive isolates were biochemically confirmed using the IMViC tests (indole, methyl red, Voges–Proskauer, and citrate) according to the method described by Feng and Weagant [12].
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed by disk diffusion on Mueller–Hinton agar (MHA; Oxoid Ltd., Basingstoke, United Kingdom) according to the Clinical and Laboratory Standards Institute (CLSI) guidelines [13]. A total of 18 antimicrobial agents were tested, including aminoglycosides (gentamicin, kanamycin, streptomycin), a beta-lactam combination (amoxicillin–clavulanic acid), cephems (cefoxitin, cefotaxime, ceftazidime, cefepime), a folate pathway antagonist (sulfamethoxazole–trimethoprim), a polymyxin (colistin), monobactams (imipenem, meropenem), a nitrofuran (nitrofurantoin), a penicillin (ampicillin), a phenicol (chloramphenicol), quinolones (ciprofloxacin, nalidixic acid), and a tetracycline (tetracycline).
E. coli American Type Culture Collection (ATCC) 25922 was used as a quality control strain. Each isolate was cultured on MHA (4 mm depth) and incubated at 37°C for 24 h. Bacterial suspensions were prepared in sterile normal saline (0.85% NaCl) and adjusted to a 0.5 McFarland standard (approximately 108 CFU/mL). The inoculum was evenly spread across MHA plates using sterile cotton swabs, antimicrobial disks were applied, and the plates were incubated at 35°C ± 2°C for 16–18 h. Inhibition zone diameters were measured with a Vernier caliper and interpreted according to CLSI (2020) criteria [13]. MDR was defined as resistance to three or more antimicrobial classes [14].
Screening for ESBL-producing isolates
ESBL-producing E. coli isolates were first identified by disk diffusion screening, in which inhibition zone diameters of ≤22 mm for ceftazidime and ≤27 mm for cefotaxime indicated potential ESBL production according to CLSI (2020) guidelines [13]. Suspected isolates were confirmed using the combination disk method with cefotaxime (30 μg) and ceftazidime (30 μg), tested alone and with clavulanic acid (10 μg) (BD, Franklin Lakes, NJ, USA). An increase of ≥5 mm in the inhibition zone diameter for the antimicrobial agent with clavulanic acid compared to the agent alone confirmed ESBL production. E. coli ATCC 25922 was used as the quality control strain.
DNA extraction
All E. coli isolates were cultured in Luria–Bertani broth (Difco, Detroit, MI, USA) at 37°C for 15–18 h. Genomic DNA was extracted using the QIAamp® DNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. DNA concentration and purity were measured with a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), and the extracted DNA was stored at −20°C until further use.
Detection of β-lactamase genes
All ESBL-producing E. coli isolates were screened for the presence of β-lactamase genes blaTEM, blaCTX-M, and blaSHV using multiplex polymerase chain reaction (PCR) assays. Primer sequences are provided in Table 1 [15–17]. Each PCR reaction (20 μL) contained Green GoTaq reaction buffer, deoxynucleoside triphosphates, forward and reverse primers, GoTaq DNA polymerase (Promega, Fitchburg, WI, USA), and template DNA. Thermocycling conditions included an initial denaturation at 94°C for 5 min, followed by 30 cycles of denaturation at 94°C for 30 s, annealing at 60°C for 30 s, and extension at 72°C for 1 min 30 s, with a final extension at 72°C for 10 min. PCR products were separated on 2.0% agarose gels stained with a red stain (Thermo Fisher Scientific, Waltham, MA, USA) and visualized under blue LED illumination using the PrepOne system (Embi Tec, San Diego, CA, USA). Reference strains carrying blaTEM, blaCTX-M, and blaSHV served as positive controls, while E. coli JM109 was used as the negative control.
Table 1. Oligonucleotide primers used for polymerase chain reaction amplification.
| Target genes | Primer | Sequence (5-3) | Amplicon Size (bp) | References |
|---|---|---|---|---|
| bla TEM | TEM-F | TCGGGGAAATGTGCG | 1074 | [15] |
| TEM-R | TGCTTAATCAGTGAGGCACC | |||
| bla SHV | SHV-F | GCCGGGTTATTCTTATTTGTCGC | 1016 | [16] |
| SHV-R | ATGCCGCCGCCAGTCA | |||
| bla CTX | CTX-M-uni-F | CGATGTGCAGTACCAGTAA | 585 | [17] |
| CTX-M-uni-R | TAAGTGACCAGAATCAGCGG | |||
| lt | LT-F | ATGACGGATATGTTTCCACTTCTC | 393 | [18] |
| LT -R | AACCTTGTGGTGCATGATGAATCC | |||
| sth | STh-F | TTCACCTTTCGCTCAGGATGCTA | 168 | [18] |
| STh-R | CACCCGGTACAAGCAGGATT | |||
| stp | STp-F | TTAATAACATCCAGCACAGGCAGG | 176 | [18] |
| STp-R | TCCCCTCTTTTAGTCAGTCAACTG | |||
| stx1A | stx1A-F2 | TCTGCAATAGGTACTCCATTACAG | 724 | [18] |
| stx1A-R2 | CCGGACACATAGAAGGAAAC | |||
| stx2A | stx2A-F2 | TTGACCATCTTCGTCTGATTATTG | 542 | [18] |
| stx2A-R2 | CTGATGATGGCAATTCAGTATAAC | |||
| aggR | aggRks | GTATACACAAAAGAAGGAAGC | 254 | [18] |
| aggRkas2 | ACAGAATCGTCAGCATCAGC | |||
| pCVD432 | CVD/1 | CTCTGGCGAAAGACTGTATC | 463 | [18] |
| CVD/2 | CATCTCTACATCAAGAGCAG | |||
| bfpA | bfpA-F | AGTCGCAGAATGCTATTTCAGAAG | 322 | [19] |
| bfpA-R | TTTTCGCCAGAGATATTAACACCG | |||
| eaeA | eaeA/1a | GCGATTACGCGAAAGATACC | 677 | [19] |
| eaeA/2a | GATAACGGAACTGCATTGAGT | |||
| ipaH | ipaH/1 | CTGGCTGATGCCGTGACAG | 801 | [19] |
| ipaH/2 | GCTGTTCAGTCTCACGCATC |
Detection of virulence genes
All 498 E. coli isolates were tested for 10 virulence genes using multiplex PCR. Primers were arranged into two sets. Primer set A targeted ETEC (lt, sth, and stp), enterohemorrhagic E. coli (EHEC) (stx1A and stx2A), and enteroaggregative E. coli (EAEC) (aggR and pCVD432) [18]. Primer set B targeted EPEC (bfpA and eaeA) and enteroinvasive E. coli (EIEC) (ipaH) [19]. Each PCR reaction (25 μL) included Green GoTaq reaction buffer, deoxynucleoside triphosphates, primers, GoTaq DNA polymerase, and template DNA. Amplification conditions featured an initial denaturation at 95°C for 5 min, followed by 35 cycles of denaturation at 95°C for 1 min, annealing at 52°C for 1 min, and extension at 72°C for 1 min, ending with a final extension at 72°C for 5 min. Amplicons were analyzed as described above. Reference strains for each virulence gene served as positive controls, and E. coli JM109 was used as a negative control.
Statistical analysis
Descriptive statistics were used to summarize the frequency and percentage of AMR, ESBL production, and virulence genes among E. coli isolates from slaughterhouses and fresh markets. Pearson’s chi-square test and Fisher’s exact test (for expected cell counts <5) were applied to compare detection rates between sample sources, and 95% confidence intervals (CIs) were calculated. Statistical analyses were conducted using the Statistical Package for the Social Sciences (SPSS version 18.0; SPSS Inc., Chicago, IL, USA). A p-value <0.05 was considered statistically significant.
RESULTS
AMR phenotypes
The antimicrobial susceptibility of all E. coli isolates was assessed across 11 antimicrobial classes using the disk diffusion method with 18 antimicrobial agents. Overall, 97.4% (485/498) of the isolates showed resistance to at least one antimicrobial agent (Table 2). Resistance was detected in 98.3% (232/236) of isolates from slaughterhouses and 96.6% (253/262) from fresh markets. No statistically significant difference in resistance rates was observed between slaughterhouse and fresh-market isolates (p > 0.05; 95% CI: 0.6–6.8).
Over 50% of E. coli isolates from slaughterhouse samples (feces and carcasses) showed resistance to ampicillin, tetracycline, chloramphenicol, streptomycin, and sulfamethoxazole–trimethoprim. Likewise, isolates from pork and cutting boards in fresh markets exhibited resistance rates over 50% for ampicillin, tetracycline, streptomycin, and chloramphenicol. Conversely, most isolates from both slaughterhouses and fresh markets remained susceptible to imipenem, meropenem, cefoxitin, and colistin.
Table 2. AMR detection rate of 498 Escherichia coli isolates recovered from slaughterhouses and fresh markets in central, Thailand.
| Antimicrobial class | Antimicrobial agents | No. (%) of AMR isolates | |||
|---|---|---|---|---|---|
|
| |||||
| Slaughterhouses | Fresh markets | ||||
|
|
| ||||
| Feces (n = 117) | Carcass (n = 119) | Pork (n = 116) | Cutting board (n = 146) | ||
| Resistance | 117 (100) | 115 (96.6) | 116 (100) | 137 (93.8) | |
| MDR | 110 (94.0) | 103 (86.6) | 107 (92.2) | 115 (78.8) | |
| Aminoglycosides | Gentamicin | 43 (36.8) | 21 (17.6) | 29 (25) | 21 (14.4) |
| Kanamycin | 36 (30.8) | 28 (23.5) | 17 (14.7) | 17 (11.6) | |
| Streptomycin | 71 (60.7) | 72 (60.5) | 73 (62.9) | 76 (52.1) | |
| Phenicols | Chloramphenicol | 80 (68.4) | 75 (63) | 67 (57.8) | 73 (50) |
| Monobactams | Imipenem | 1 (0.9) | 3 (2.5) | 7 (6.0) | 3 (2.1) |
| Meropenem | 0 | 3 (2.5) | 0 | 1 (0.7) | |
| Cephems | Cefoxitin | 1 (0.9) | 3 (2.5) | 4 (3.4) | 3 (2.1) |
| Cefotaxime | 41 (35) | 28 (23.5) | 27 (23.3) | 20 (13.7) | |
| Ceftazidime | 13 (11.1) | 6 (5.0) | 6 (5.2) | 8 (5.5) | |
| Cefepime | 27 (23.1) | 18 (15.1) | 14 (12.1) | 12 (8.2) | |
| Nitrofurans | Nitrofuratoin | 7 (6.0) | 5 (4.2) | 10 (8.6) | 3 (2.1) |
| Penicillins | Ampicillin | 112 (95.7) | 109 (91.6) | 115 (99.1) | 132 (90.4) |
| β-lactam combination | Amoxicillin/clavulanic acid | 11 (9.4) | 16 (13.4) | 18 (15.5) | 0 |
| Polymyxins | Colistin | 2 (1.7) | 9 (7.6) | 7 (6) | 5 (3.4) |
| Quinolones | Ciprofloxacin | 25 (21.4) | 26 (21.8) | 19 (16.4) | 10 (6.8) |
| Nalidixic acid | 52 (44.4) | 41 (34.5) | 38 (32.8) | 30 (20.5) | |
| Folate pathway antagonists | Sulfamethoxazole/ Trimethoprim | 62 (53) | 66 (55.5) | 56 (48.3) | 69 (47.3) |
| Tetracyclines | Tetracycline | 98 (83.8) | 95 (79.8) | 98 (84.5) | 100 (68.5) |
AMR = Antimicrobial resistance, MDR = Multidrug resistance.
MDR and AMR patterns
All 498 E. coli isolates were evaluated for MDR based on resistance to various antimicrobial classes. A high rate of MDR was observed, with 87.3% (435/498) of isolates identified as MDR (Table 2). MDR rates were high in both slaughterhouse (90.3%; 213/236) and fresh-market (84.7%; 222/262) isolates. There was no statistically significant difference in MDR prevalence between the two sources (p > 0.05; 95% CI: 0.9–2.8).
A total of 130 unique AMR patterns were identified among all isolates (Supplementary Table 1). The most common patterns were AMI–PHE–PEN–FOL–TET (50 isolates), AMI–PEN–TET (24 isolates), and AMI–PHE–PEN–QUI–FOL–TET (24 isolates).
Prevalence of ESBL-producing E. coli
ESBL-producing E. coli accounted for 23.5% (117/498) of all isolates. Of these, 14.3% (71/498) came from slaughterhouses, while 9.2% (46/498) were recovered from fresh markets. The prevalence of ESBL-producing isolates was significantly higher in slaughterhouses than in fresh markets (p < 0.05; 95% CI: 1.3–3.1).
Among slaughterhouse samples, ESBL-producing E. coli were found in 34.2% (40/117) of fecal isolates and 26.1% (31/119) of carcass isolates. In fresh markets, ESBL-producing isolates appeared in 18.9% (23/116) of pork samples and 15.8% (23/146) of cutting board samples (Table 3).
Table 3. Detection rate of β-lactamase genes in 117 ESBL-producing Escherichia coli isolates from slaughterhouses and fresh markets in central Thailand.
| Sample types | ESBL-producing E. coli (%) | No. (%) of E. coli harboring the β-lactamase gene | ||||
|---|---|---|---|---|---|---|
|
| ||||||
| bla TEM | bla CTX-M | bla SHV | blaTEM + blaCTX-M | Not determined | ||
| Slaughterhouses | 71 (14.3) | 49 (20.8) | 54 (22.9) | 0 | 34 (14.4) | 0 |
| Feces (n=117) | 40 | 25 (62.5) | 34 (85) | 0 | 18 (45) | 0 |
| Carcass (n=119) | 31 | 24 (77.4) | 20 (64.5) | 0 | 16 (51.6) | 3 (9.7) |
| Fresh markets | 46 (9.2) | 29 (11.1) | 15 (5.7) | 2 (0.8) | 8 (3.1) | 9 (19.6) |
| Pork (n=116) | 23 | 13 (56.5) | 5 (21.7) | 0 | 2 (8.7) | 7 (30.4) |
| Cutting board (n = 146) | 23 | 16 (69.6) | 10 (43.5) | 2 (8.7) | 6 (26.1) | 2 (8.7) |
| Total | 117 (23.5) | 78 (66.7) | 69 (59) | 2 (1.7) | 42 (35.9) | 12 (10.3) |
E. coli = Escherichia coli, ESBL = Extended-spectrum β-lactamase.
* Significantly different (χ2; p < 0.05) *Not determined (none of detected genes).
MDR among ESBL-producing isolates
Among the 117 ESBL-producing E. coli isolates, 97.4% (114/117) were classified as MDR. The prevalence of MDR was similarly high in isolates from slaughterhouses (97.2%; 69/71) and fresh markets (97.8%; 45/46). Among slaughterhouse-derived ESBL producers, MDR was found in 34.2% (40/117) of fecal isolates and 24.8% (29/117) of carcass isolates. In fresh markets, MDR ESBL-producing isolates were present in 19.7% (23/117) of pork samples and 18.8% (22/117) of cutting board samples. Three ESBL-producing isolates were not classified as MDR.
A total of 44 unique AMR patterns were identified among ESBL-producing isolates. The most common patterns were AMI–PHE–CEP–PEN–FOL–TET (16 isolates) and AMI–PHE–CEP–PEN–QUI–FOL–TET (16 isolates) (Figure 1).
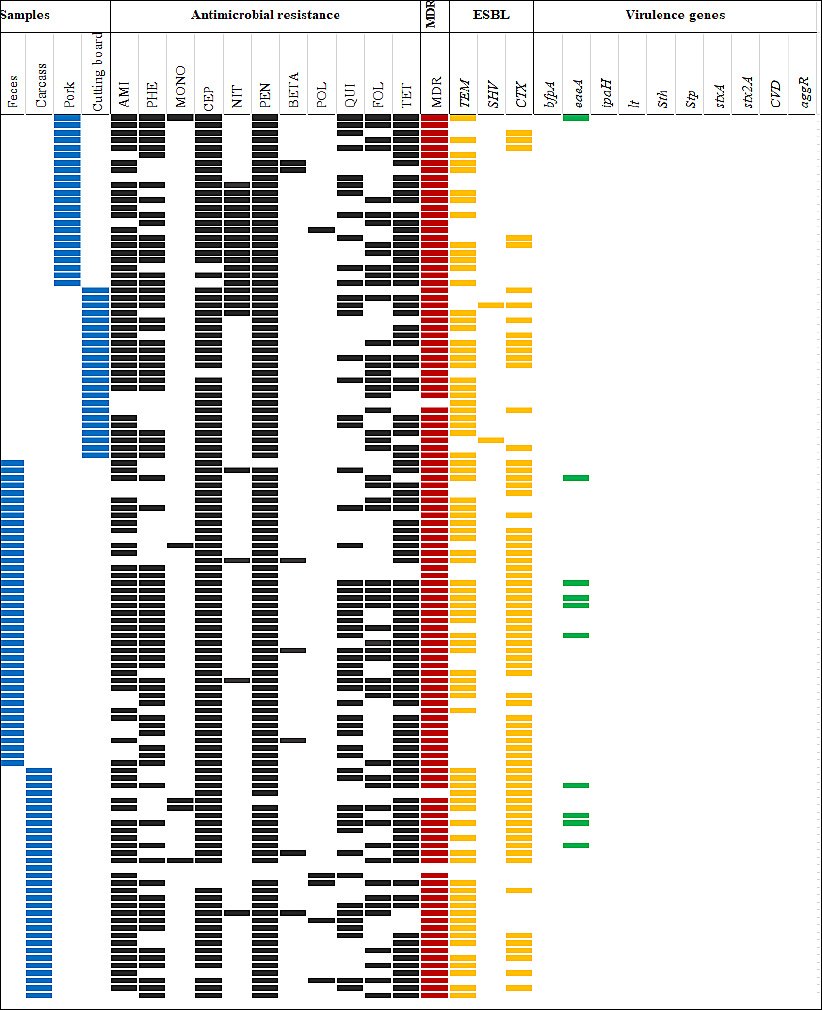
Figure 1. Antimicrobial resistance (AMR) patterns, multidrug resistance (MDR), virulence genes, and β-lactamase genes of 117 ESBL-producing Escherichia coli isolates recovered from slaughterhouses and fresh markets in central Thailand. The solid boxes indicate a feature in the isolate. Blue boxes indicate sample types, black boxes indicate AMR, red boxes indicate MDR, yellow boxes indicate ESBL genes, and green boxes indicate virulence genes. AMI = Aminoglycosides, BETA = β-lactam combination, CEP = Cephems, FOL = Folate pathway antagonists, MONO = Monobactams, NIT = Nitrofurans, PEN = Penicillins, PHE = Phenicols, POL = Polymyxins, QUI = Quinolones, TET = Tetracyclines.
Distribution of β-lactamase genes in ESBL-producing isolates
Among the 117 ESBL-producing E. coli isolates, 89.7% (105/117) carried at least one of the tested β-lactamase genes (blaTEM, blaCTX-M, or blaSHV). The blaTEM gene was the most commonly detected, found in 66.7% (78/117) of isolates, including 69.0% (49/71) from slaughterhouses and 63.0% (29/46) from fresh markets. The blaCTX-M gene was present in 59.0% (69/117) of isolates, with a significantly higher prevalence in slaughterhouse isolates (76.0%; 54/71) compared to those from fresh markets (32.6%; 15/46).
The blaSHV gene was detected at a low frequency (4.3%; 2/46) and was found only in isolates from fresh markets. Co-occurrence of blaTEM and blaCTX-M was observed in 35.9% (42/117) of isolates, including 47.9% (34/71) from slaughterhouses and 17.4% (8/46) from fresh markets. The prevalence of blaTEM, blaCTX-M, and their combination was significantly higher in slaughterhouse isolates than in fresh-market isolates (p < 0.05) (Table 3).
Detection of virulence genes in E. coli isolates
Virulence genes were found in 3.2% (16/498) of E. coli isolates, indicating the presence of pathogenic strains. Three virulence genes, eaeA, lt, and stp, were identified (Table 4). The eaeA gene, which encodes intimin and is linked to EPEC and EHEC, was detected in 4.7% (11/236) of isolates from slaughterhouses and 1.1% (3/262) from fresh markets. The lt and stp genes were each found at a low frequency of 0.8% (2/262) in fresh-market samples, with lt identified in one cutting board isolate and stp in one pork isolate.
No statistically significant difference was observed in the prevalence of pathogenic E. coli between slaughterhouse and fresh-market isolates (p > 0.05; 95% CI: 0.8–7.3). Of the 16 virulence gene–positive isolates, 62.5% (10/16) were MDR and carried β-lactamase genes. The virulence genes bfpA, ipaH, sth, stx1A, stx2A, aggR, and pCVD432 were not detected in any isolate.
Table 4. Detection rate of virulence genes in 498 Escherichia coli isolates recovered from slaughterhouses and fresh markets in central Thailand.
| Pathotype | Virulence genes | Number (%) of virulence genes | |||
|---|---|---|---|---|---|
|
| |||||
| Slaughterhouses | Fresh markets | ||||
|
|
| ||||
| Feces (n=117) | Carcass (n=119) | Pork (n=116) | Cutting board (n=146) | ||
| EPEC/EHEC | eaeA | 5 (4.3) | 6 (5.0) | 3 (2.6) | 0 |
| ETEC | It | 0 | 0 | 0 | 1 (0.7) |
| stp | 0 | 0 | 1 (0.9) | 0 | |
| Total | 5 (4.3) | 6 (5.0) | 4 (3.4) | 1 (0.7) | |
EPEC = Enteropathogenic Escherichia coli, EHEC= Enterohemorrhagic E. coli, ETEC = Enterotoxigenic E. coli
DISCUSSION
AMR patterns along the pork production chain
Antimicrobial agents are commonly used in the pork production chain, especially at the farm-level for therapeutic, growth-promoting, and disease-preventing purposes [20]. However, such practices promote the emergence and persistence of AMR at various stages of pork production. In this study, more than 50% of E. coli isolates from slaughterhouse samples (feces and carcasses) and fresh-market samples (pork and cutting boards) showed resistance to ampicillin, tetracycline, chloramphenicol, and streptomycin. These antimicrobials are among the ones most frequently linked with resistance in E. coli isolates from pigs [21].
The observed resistance patterns align with earlier reports from pig farms in Thailand [4, 22] and slaughterhouse-based studies in the Mekong Delta, Vietnam [23], Brazil [24], and Australia [25]. Notably, studies from Brazil and Australia reported high resistance rates to ampicillin (81.1% and 60.2%, respectively) and tetracycline (97.8% and 68.2%, respectively), along with significant resistance to chloramphenicol [24, 25]. High volumes of tetracyclines, penicillins, and sulfonamides sold for veterinary use have been documented [26], and the improper or excessive use of these antimicrobials in pig production likely drives the elevated resistance rates observed [27]. The detection of chloramphenicol-resistant isolates is especially concerning, as this antimicrobial has been banned for use in food-producing animals [28]. Such resistance may persist due to co-selection or cross-resistance caused by ongoing use of other antimicrobials [29], emphasizing the complex and multifactorial nature of AMR development [28, 30].
Colistin resistance was found in 4.6% of isolates from both slaughterhouse and fresh-market samples. Since colistin is considered a last-resort antibiotic for treating MDR gram-negative infections, this finding raises serious public health concerns. The plasmid-mediated mcr-1 gene is key in spreading colistin resistance among animal, environmental, and human reservoirs, enabling rapid horizontal transfer [31].
Occurrence and distribution of ESBL-producing E. coli
In this study, 23.5% of E. coli isolates were identified as ESBL producers, a prevalence similar to that previously reported in healthy pigs in Thailand (19.2%) [4]. The detection rate of ESBL-producing E. coli was significantly higher in slaughterhouses (14.3%) than in fresh markets (9.2%) (p < 0.05), indicating greater contamination pressure at earlier stages of the pork production process rather than later stages.
Nevertheless, ESBL prevalence varied greatly when compared to other studies conducted in Thailand and other countries. For example, Boonyasiri et al. [32] reported ESBL prevalence of 33.3% in slaughterhouses and 61.5% in markets in eastern and northern Thailand, while Sornsenee et al. [33] documented ESBL-producing E. coli in minced chicken (79.17%), pork (43.75%), and beef (22.73%) in southern Thailand. Studies from other nations have also shown significant ESBL contamination in food animals and retail meat, including Singapore [34], Vietnam [35], South Korea [36], and Cameroon [37]. Such differences likely result from variations in sample types, sampling strategies, timing, farming practices, slaughterhouse hygiene, and antimicrobial use patterns.
The detection of ESBL-producing E. coli in both slaughterhouse and fresh-market samples indicates potential risks of transmission to consumers and environmental spread. Maintaining strict hygiene practices during and after slaughter is therefore essential to reduce fecal contamination and limit ESBL dissemination [33]. The presence of ESBL-producing E. coli on pork and cutting boards further suggests cross-contamination, likely from fecal sources, although molecular typing is needed to confirm transmission pathways.
MDR among ESBL-producing isolates
An extremely high proportion (97.4%) of ESBL-producing E. coli isolates showed MDR, highlighting the serious clinical and public health concerns of ESBL phenotypes in pork production systems. Similar MDR rates have been reported in Thailand [33, 38], Vietnam [35], and Cameroon [37]. The high MDR frequency may result from extensive antimicrobial use in industrial pig farming, especially β-lactam antimicrobials used for disease prevention and growth promotion [39]. Such practices exert strong selective pressure, fostering the emergence and persistence of MDR organisms capable of spreading through food production systems to humans and the environment [40].
β-lactamase gene profiles and their epidemiological significance
β-lactamase genes were found at significantly higher rates in ESBL-producing E. coli isolates from slaughterhouses compared to those from fresh markets (p < 0.05), indicating that slaughterhouses are a key control point for ESBL spread. Overall, 89.7% of ESBL-producing isolates carried at least one bla gene, with blaTEM being the most common (66.7%). The TEM enzyme is the most widespread β-lactamase among gram-negative bacteria, providing resistance to penicillins and early-generation cephalosporins commonly used in pig farming [41, 42]. Similar high prevalence of blaTEM has been reported in Thailand among healthy pigs and minced meat [33, 43].
The blaCTX-M gene was found in 59% of ESBL-producing isolates, aligning with previous reports from Thailand [44] and other countries, including the United Kingdom, Germany, Tunisia, and Switzerland [45]. Notably, 35.9% of isolates carried more than one bla gene, providing evidence that ESBL-associated plasmids often contain multiple resistance determinants [46, 47]. These genetic configurations likely help maintain and spread β-lactam resistance throughout the pork production chain.
Virulence gene distribution in E. coli isolates
Virulence genes were detected in only 3.2% of E. coli isolates, indicating a low prevalence of pathogenic E. coli within the study group. The identified virulence genes included eaeA, lt, and stp. The eaeA gene, which encodes intimin and is linked to EPEC and EHEC, was detected in both slaughterhouse and fresh-market samples and is associated with diarrheal illness in humans. The lt and stp genes, responsible for producing heat-labile and heat-stable enterotoxins in ETEC, respectively, were detected at very low levels [8, 48].
The low detection rate of virulence genes may be due to the age of the slaughtered pigs or to the lack of active infection at the time of sampling. Notably, the contrasting pattern seen in this study, high prevalence of AMR and ESBL production but low virulence gene detection, indicates that the selective pressures promoting AMR are different from those affecting the distribution of genes associated with pathogenicity.
Limitations
This study has several limitations. First, using archived isolates collected during 2017–2018 limited the ability to directly trace or confirm cross-contamination pathways between slaughterhouses and individual fresh markets. Second, only a limited set of virulence genes was examined, which may underestimate the overall pathogenic potential of circulating E. coli strains. Third, molecular typing methods, such as whole-genome sequencing, were not performed due to financial constraints, thereby restricting the assessment of genetic relatedness and transmission dynamics along the pork production chain. Finally, environmental samples were not included, preventing the identification of potential AMR reservoirs within slaughterhouses and fresh markets. Despite these limitations, the study provides valuable baseline data to inform future research and guide targeted interventions to reduce AMR dissemination in pork production systems.
CONCLUSION
This study revealed a significant burden of AMR throughout the pork production chain in central Thailand. Nearly all E. coli isolates showed resistance to at least one antimicrobial agent, with a large proportion classified as MDR. ESBL-producing E. coli were found frequently, with a notably higher prevalence in slaughterhouse samples compared to fresh-market samples, underscoring slaughterhouses as key upstream contamination sources. Most ESBL-producing isolates carried β-lactamase genes, mainly blaTEM and blaCTX-M, and almost all exhibited MDR phenotypes. Conversely, virulence genes were rarely detected, suggesting that resistance traits are more widespread than pathogenic factors among the circulating E. coli populations.
The findings highlight the importance of slaughterhouses and fresh markets as key points in the spread of antimicrobial-resistant E. coli in the pork production chain. These settings allow fecal contamination, cross-contamination of carcasses, meat, and food-contact surfaces, and exposure of consumers. Improving hygiene and sanitation practices during slaughter, carcass processing, and retail handling is vital to decreasing the spread of resistant bacteria. Moreover, the high prevalence of MDR and ESBL-producing E. coli underscores the urgent need for cautious antimicrobial use in pig farming, especially by restricting the use of critically important antimicrobials. Coordinated surveillance across farms, slaughterhouses, and markets is essential to support evidence-based actions within a One Health approach.
A key strength of this study is the comprehensive assessment of AMR phenotypes, ESBL production, β-lactamase gene distribution, and virulence profiles in E. coli isolates collected from different stages of the pork production process. Including both slaughterhouse and fresh-market samples enabled direct comparison of upstream and downstream contamination points. Additionally, the relatively large number of isolates analyzed provides a solid baseline for understanding AMR dynamics in a region with intensive pig production.
Future research should include longitudinal sampling and environmental monitoring to better understand how antimicrobial-resistant E. coli spreads along the pork production process. Using molecular typing methods, such as whole-genome sequencing, would allow detailed analysis of strain relationships, resistance gene transfer, and potential contamination sources. Broadening the scope of virulence genes studied and adding data on antimicrobial use at the farm-level would enhance risk assessment and help develop targeted mitigation strategies.
In conclusion, this study emphasizes the widespread occurrence of AMR, MDR, and ESBL-producing E. coli throughout the pork production chain in central Thailand, with slaughterhouses identified as key points for the spread of resistance. Although pathogenic E. coli were found at low levels, the widespread presence of resistance factors presents a serious public health issue. Coordinated actions targeting antimicrobial stewardship, better hygiene practices, and ongoing surveillance are vital to prevent the transfer of AMR from pork production to humans and the environment.
DATA AVAILABILITY
The supplementary data can be available from the corresponding author.
AUTHORS’ CONTRIBUTIONS
OS and WP: Designed the study, collected samples, analyzed data, and drafted and revised the manuscript. YZ and CN: Designed the study, drafted, and revised the manuscript. AS, SW, AI, and RP: Analyzed data and drafted and revised the manuscript. All authors have read and approved the final version.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the partial financial support provided by the Thammasat University Research Fund under a TU Research Scholar contract (8/2560) awarded to O. Suthienkul. This work was supported in part by the Japan Agency for Medical Research and Development (AMED) under Grant Numbers JP20wm0125008 and JP223fa627005 to Y. Suzuki. Additionally, this research received partial support from the Faculty of Veterinary Medicine, Mahanakorn University of Technology, Thailand.
REFERENCES
- Mancuso G, Midiri A, Gerace E, Biondo C. Bacterial antibiotic resistance:The most critical pathogens. Pathogens (Basel) 2021;10(10):1310. [Google Scholar] | [Crossref]
- Chonsin K, Changkwanyeun R, Siriphap A, Intarapuk A, Prapasawat W, Changkaew K, Pulsrikarn C, Isoda N, Nakajima C, Suzuki Y, Suthienkul O. Prevalence and multidrug resistance of Salmonella in swine production chain in a central province, Thailand. J Food Prot 2021;84(12):2174-2184. [Google Scholar] | [Crossref]
- Lekagul A, Tangcharoensathien V, Liverani M, Mills A, Rushton J, Yeung S. Understanding antibiotic use for pig farming in Thailand:A qualitative study. Antimicrob Resist Infect Control 2021;10(1):3. [Google Scholar] | [Crossref]
- Trongjit S, Assavacheep P, Samngamnim S, My TH, An VTT, Simjee S, Chuanchuen R. Plasmid-mediated colistin resistance and ESBL production in Escherichia coli from clinically healthy and sick pigs. Sci Rep 2022;12(1):2466. [Google Scholar] | [Crossref]
- Ma F, Xu S, Tang Z, Li Z, Zhang L. Use of antimicrobials in food animals and impact of transmission of antimicrobial resistance on humans. Biosaf Health 2021;3(1):32-38. [Google Scholar] | [Crossref]
- Castanheira M, Simner PJ, Bradford PA. Extended-spectrum beta-lactamases:An update on their characteristics, epidemiology and detection. JAC Antimicrob Resist 2021;3(3):dlab092. [Google Scholar] | [Crossref]
- Songsaeng W, Am-in N, Prapasarakul N, Sirichokchatchawan W. Multidrug-resistant ESBL-producing Escherichia coli coexisting with colistin-resistance genes in pig farms, Central Thailand. Thai J Vet Med 2024;54(1):69-76. [Google Scholar] | [Crossref]
- Barros MM, Castro J, Araújo D, Campos AM, Oliveira R, Silva S, Outor-Monteiro D, Almeida C. Swine colibacillosis:Global epidemiologic and antimicrobial scenario. Antibiotics (Basel) 2023;12(4):682. [Google Scholar] | [Crossref]
- Do KH, Byun JW, Lee WK. Antimicrobial resistance, adhesin and toxin genes of porcine pathogenic Escherichia coli following the ban on antibiotics as the growth promoters in feed. Pak Vet J 2021;41(4):519-523. [Google Scholar] | [Crossref]
- Yang SC, Lin CH, Aljuffali IA, Fang JY. Current pathogenic Escherichia coli foodborne outbreak cases and therapy development. Arch Microbiol 2017;199(6):811-825. [Google Scholar] | [Crossref]
- Agriculture in Thailand. Bangkok: Office of Agricultural Economics; 1992. [Google Scholar]
- Feng P, Weagant SD. BAM Chapter 4A:Diarrheagenic Escherichia coli. Silver Spring: US Food and Drug Administration; 2020. [Google Scholar]
- Performance standards for antimicrobial susceptibility testing;30th informational supplement. M100. Wayne (PA): CLSI; 2020. [Google Scholar]
- Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG, Harbarth S, Hindler JF, Kahlmeter G, Olsson-Liljequist B, Paterson DL. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria:An international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect 2012;18(3):268-281. [Google Scholar] | [Crossref]
- Yu Y, Ji S, Chen Y, Zhou W, Wei Z, Li L, Ma Y. Resistance of strains producing extended-spectrum β-lactamases and genotype distribution in China. J Infect 2007;54(1):53-57. [Google Scholar] | [Crossref]
- Rayamajhi N, Kang SG, Lee DY, Kang ML, Lee SI, Park KY, Lee HS, Yoo HS. Characterization of TEM, SHV and AmpC-type β-lactamases from cephalosporin-resistant Enterobacteriaceae isolated from swine. Int J Food Microbiol 2008;124(2):183-187. [Google Scholar] | [Crossref]
- Batchelor M, Hopkins K, Threlfall EJ, Clifton-Hadley FA, Stallwood AD, Davies RH, Liebana E. blaCTX-M genes in clinical Salmonella isolates recovered from humans in England and Wales from, 1992 to 2003. Antimicrob Agents Chemother 2005;49(4):1319-1322. [Google Scholar] | [Crossref]
- Prapasawat W, Intarapuk A, Chompook P, Nakajima C, Suzuki Y, Suthienkul O. Antimicrobial resistance, integron, virulence gene, and multilocus sequence typing of Escherichia coli isolates from postweaning piglets with and without diarrhea. Southeast Asian J Trop Med Public Health 2017;48(5):1042-1055. [Google Scholar] | [Crossref]
- Pachanon R. Detection of pathogenic Escherichia coli and their antimicrobial resistance in diarrheal swine [master's thesis]. Bangkok: Mahidol University; 2014. [Google Scholar]
- Ferraz MP. Antimicrobial resistance:The impact from and on society according to One Health approach. Soc 2024;14(9):187. [Google Scholar] | [Crossref]
- Pazra DF, Iryawati D. Genetic characteristics of antimicrobial resistance in Escherichia coli isolated from farm animals, slaughterhouses, and associated environments. London: IntechOpen; 2025. [Google Scholar]
- Prapasarakul N, Tummaruk P, Niyomtum W, Tripipat T, Serichantalergs O. Virulence genes and antimicrobial susceptibilities of hemolytic and nonhemolytic Escherichia coli isolated from post-weaning piglets in central Thailand. J Vet Med Sci 2010;72(12):1603-1608. [Google Scholar] | [Crossref]
- Van Thong N, Thuan NK, Le Minh BT. Prevalence of antibiotic resistance genes of Escherichia coli at the pig slaughterhouses in the Mekong Delta. Vet Integr Sci 2024;23(1):1-10. [Google Scholar] | [Crossref]
- Pissetti C, Werlang GO, Kich JD, Cardoso M. Genotyping and antimicrobial resistance in Escherichia coli from pig carcasses. Pesq Vet Bras 2017;37:1253-1260. [Google Scholar] | [Crossref]
- Kidsley AK, Abraham S, Bell JM, O'Dea M, Laird TJ, Jordan D, Mitchell P, McDevitt CA, Trott DJ. Antimicrobial susceptibility of Escherichia coli and Salmonella spp isolates from healthy pigs in Australia:Results of a pilot national survey. Front Microbiol 2018;9:1207. [Google Scholar] | [Crossref]
- European surveillance of veterinary antimicrobial consumption, 2022. Sales of veterinary antimicrobial agents in 31 European countries in 2022. Amsterdam: EMA; 2023. [Google Scholar]
- Gonçalves C, Silveira L, Rodrigues J, Furtado R, Ramos S, Nunes A, Pista Â. Phenotypic and genotypic characterization of Escherichia coli and Salmonella spp, isolates from pigs at slaughterhouse and from commercial pork meat in Portugal. Antibiotics (Basel) 2024;13(10):957. [Google Scholar] | [Crossref]
- Trongjit S, Angkittitrakul S, Chuanchuen R. Occurrence and molecular characteristics of antimicrobial resistance of Escherichia coli from broilers, pigs and meat products in Thailand and Cambodia provinces. J Microbiol Immunol 2016;60(9):575-585. [Google Scholar] | [Crossref]
- Sheikh M, Gholipour S, Ghodsi S, Nikaeen M. Co-selection of antibiotic and disinfectant resistance in environmental bacteria:Health implications and mitigation strategies. Environ Res 2025;267:120708. [Google Scholar] | [Crossref]
- Hossain AZ, Chowdhury AMA. Understanding the evolution and transmission dynamics of antibiotic resistance genes:A comprehensive review. J Basic Microbiol 2024;64(10):e2400259. [Google Scholar] | [Crossref]
- Treilles M, Châtre P, Drapeau A, Madec JY, Haenni M. Spread of the mcr-1 colistin-resistance gene in Escherichia coli through plasmid transmission and chromosomal transposition in French goats. Front Microbiol 2023;13:1023403. [Google Scholar] | [Crossref]
- Boonyasiri A, Tangkoskul T, Seenama C, Saiyarin J, Tiengrim S, Thamlikitkul V. Prevalence of antibiotic resistant bacteria in healthy adults, foods, food animals, and the environment in selected areas in Thailand. Pathog Glob Health 2014;108(5):235-245. [Google Scholar] | [Crossref]
- Sornsenee P, Chimplee S, Arbubaker A, Kongchai S, Madimong H, Romyasamit C. Occurrence, antimicrobial resistance profile, and characterization of extended-spectrum β-lactamase-producing Escherichia coli isolates from minced meat at local markets in Thailand. Foodborne Pathog Dis 2022;19(3):232-240. [Google Scholar] | [Crossref]
- Guo S, Aung KT, Leekitcharoenphon P, Tay MY, Seow KL, Zhong Y, Ng LC, Aarestrup FM, Schlundt J. Prevalence and genomic analysis of ESBL-producing Escherichia coli in retail raw meats in Singapore. J Antimicrob Chemother 2021;76(3):601-605. [Google Scholar] | [Crossref]
- Le QP, Ueda S, Nguyen TNH, Dao TVK, Van Hoang TA, Tran TTN, Hirai I, Nakayama T, Kawahara R, Do TH, Vien QM. Characteristics of extended-spectrum β-lactamase–producing Escherichia coli in retail meats and shrimp at a local market in Vietnam. Foodborne Pathog Dis 2015;12(8):719-725. [Google Scholar] | [Crossref]
- Kim YJ, Moon JS, Oh DH, Chon JW, Song BR, Lim JS, Heo EJ, Park HJ, Wee SH, Sung K. Genotypic characterization of ESBL-producing Escherichia coli from imported meat in South Korea. Food Res Int 2018;107:158-164. [Google Scholar] | [Crossref]
- Matakone M, Founou RC, Founou LL, Dimani BD, Koudoum PL, Fonkoua MC, Boum-Ii Y, Gonsu H, Noubom M. Multidrug-resistant and extended-spectrum β-lactamase-producing Escherichia coli isolated from slaughtered pigs and slaughterhouse workers in Yaoundé, Cameroon. One Health 2024;19:100885. [Google Scholar] | [Crossref]
- Saenkankam I, Apiwatsiri P, Supimon N, Niyomtham W, Hampson DJ, Prapasarakul N. Tracking extended-spectrum β-lactamase-producing and colistin-resistant Escherichia coli in pig abattoirs:Impacts on food safety. Foodborne Pathog Dis 2025;10(1089):40229124. [Google Scholar] | [Crossref]
- Coelho MMS, Davanzo EFA, Dos Santos RL, Castro VHDL, da Costa HMB, Dallago BSL, Perecmanis S, Santana AP. Escherichia coli and Enterobacteriaceae counts, virulence gene profile, antimicrobial resistance, and biofilm formation capacity during pig slaughter stages. Life 2024;14(10):1261. [Google Scholar] | [Crossref]
- Hide M, Meng S, Cheng S, Bañuls AL, Ky S, Yay C, Laurent D, Delvallez G. Colistin resistance in ESBL- and carbapenemase-producing Escherichia coli and Klebsiella pneumoniae clinical isolates in Cambodia. J Glob Antimicrob Resist 2024;38:236-244. [Google Scholar] | [Crossref]
- Hussain HI, Aqib AI, Seleem MN, Shabbir MA, Hao H, Iqbal Z, Kulyar MFEA, Zaheer T, Li K. Genetic basis of molecular mechanisms in β-lactam resistant gram-negative bacteria. Microb Pathog 2021;158:105040. [Google Scholar] | [Crossref]
- Lekagul A, Tangcharoensathien V, Mills A, Rushton J, Yeung S. How antibiotics are used in pig farming:A mixed-methods study of pig farmers, feed mills and veterinarians in Thailand. BMJ Glob Health 2020;5(2):e001918. [Google Scholar] | [Crossref]
- Lugsomya K, Chatsuwan T, Niyomtham W, Tummaruk P, Hampson DJ, Prapasarakul N. Routine prophylactic antimicrobial use is associated with increased phenotypic and genotypic resistance in commensal Escherichia coli isolates recovered from healthy fattening pigs on farms in Thailand. Microb Drug Resist 2018;24(2):213-223. [Google Scholar] | [Crossref]
- Lay KK, Torio HE, Bitrus AA, Mala W, Sinwat N, Chuanchuen R. Multidrug-resistant Escherichia coli harboring extended-spectrum β-lactamase-encoding genes isolated from clinically healthy pigs. Thai J Vet Med 2021;51(2):303-310. [Google Scholar] | [Crossref]
- Valentin L, Sharp H, Hille K, Seibt U, Fischer J, Pfeifer Y, Michael GB, Nickel S, Schmiedel J, Falgenhauer L, Friese A. Subgrouping of ESBL-producing Escherichia coli from animal and human sources:An approach to quantify the distribution of ESBL types between different reservoirs. Int J Med Microbiol 2014;304(7):805-816. [Google Scholar] | [Crossref]
- Chishimba K, Hang'Ombe BM, Muzandu K, Mshana SE, Matee MI, Nakajima C, Suzuki Y. Detection of extended-spectrum beta-lactamase-producing Escherichia coli in market-ready chickens in Zambia. Int J Microbiol 2016;2016:5275724. [Google Scholar] | [Crossref]
- Husna A, Rahman MM, Badruzzaman ATM, Sikder MH, Islam MR, Rahman MT, Alam J, Ashour HM. Extended-spectrum β-lactamases:Challenges and opportunities. Biomedicines 2023;11(11):2937. [Google Scholar] | [Crossref]
- Ray B, Bhunia A. Escherichia coli gastroenteritis. New York: CRC Press; 2008. p. 323-326. [Google Scholar]