


ABSTRACT
Background and Aim: The fatty acid-binding protein 4 (
Materials and Methods: Blood samples were collected from 95 Bali cattle bulls sourced from a single population. Genomic DNA was extracted, and a 721-bp fragment of
Results: Four novel
Conclusion: This study provides the first evidence of
Keywords: Bali cattle,
INTRODUCTION
Meat quality and carcass traits are economically important in beef production because they directly affect processing efficiency, market value, and consumer acceptance [1]. Key attributes, including muscle development, tenderness, intramuscular fat (IMF), and fatty acid (FA) composition [2], play central roles in determining meat palatability and nutritional quality. Improvements in these traits can substantially enhance the market appeal of meat products by meeting consumer expectations for superior taste and texture while also providing health benefits through improved FA profiles [3]. Inoue
In Indonesia, Bali cattle (
Meat quality and carcass traits are complex phenotypes regulated by multiple genes. The application of DNA markers in marker-assisted selection (MAS) has markedly advanced livestock breeding [9]. Developments in molecular genetics have enabled the identification of candidate genes associated with economically important traits, thereby improving breeding efficiency through MAS [10, 11]. This approach allows researchers to detect specific genetic variations linked to desirable phenotypes. However, integrative genotype–phenotype analyses combining
The fatty acid-binding protein 4 (
Despite growing evidence that
Therefore, this study aimed to identify and characterize SNPs in the
MATERIALS AND METHODS
Ethical approval
The Animal Ethics Committee of the Banjarmasin City Food Security, Agriculture, and Fisheries Service approved the experimental procedures (approval number: 520/624/DKP3/X11/2021). All sampling procedures complied with established animal welfare guidelines. Gentle restraint, appropriately sized needles, and rapid venipuncture performed by trained personnel were used to minimize pain and stress during blood collection. The puncture site was cleaned before and after sampling to reduce discomfort and promote healing. Slaughter was conducted at a licensed slaughterhouse in accordance with Indonesian regulations for humane handling and slaughter of cattle.
Study period and location
The experiment was conducted from December 2022 to June 2023. A total of 95 Bali cattle bulls from a single population were included, and all available animals were used as representative samples. The animals weighed 250–350 kg and were 18–36 months of age. Bali cattle were sourced from Kupang, East Nusa Tenggara Province, Indonesia, and transported by ship to the Basirih slaughterhouse in Banjarmasin. Upon arrival, cattle were maintained under an intensive management system for 2 weeks before slaughter in South Banjarmasin, South Kalimantan. During this period, forage was provided at 10% of body weight, concentrate at 2%, and water
Blood collection and DNA extraction
Blood samples were collected from the jugular vein using a Venoject with a sterile disposable 21-gauge needle (Becton, Dickinson and Company, Franklin Lakes, NJ, USA). Approximately 5 mL of blood was transferred into vacuum tubes containing 1.5 mL ethylenediaminetetraacetic acid (EDTA). Samples were immediately placed on ice and stored at 4°C until DNA extraction. Genomic DNA was extracted using the Geneaid Genomic DNA Mini Kit (Geneaid Biotech Ltd., Taipei, Taiwan) according to the manufacturer’s instructions. Extracted DNA was stored at −20°C. DNA purity was assessed spectrophotometrically, with acceptable A260/280 ratios of 1.8–2.2 and concentrations ≥20 ng/μL.
Polymerase chain reaction (PCR) amplification of FABP4
Primer sequences were designed using Primer3 and BLAST primer tools. The forward (5′-CCC TCC ATC ATT GTA ATC ACT-3′) and reverse (5′-GGA CAA CGT ATC CAG CAG AAA-3′) primers for the
Agarose gel electrophoresis and DNA sequencing
PCR products were separated on a 1% agarose gel stained with Florosafe (1st BASE, Singapore, Singapore) and visualized using a UV transilluminator (Bio-Rad, Hercules, CA, USA). Sequencing was performed at 1st BASE Laboratory Services (Selangor, Malaysia) using an ABI PRISM system with the BigDye Terminator kit v3.1 (Applied Biosystems). Sequencing quality was manually examined using FinchTV, BioEdit, and Molecular Evolutionary Genetic Analysis software. The first and last 30 bp were excluded because of low peak quality, and only SNPs with clear chromatogram peaks were retained. Sequence alignment was conducted against the Ensembl
Ultrasound imaging measurements
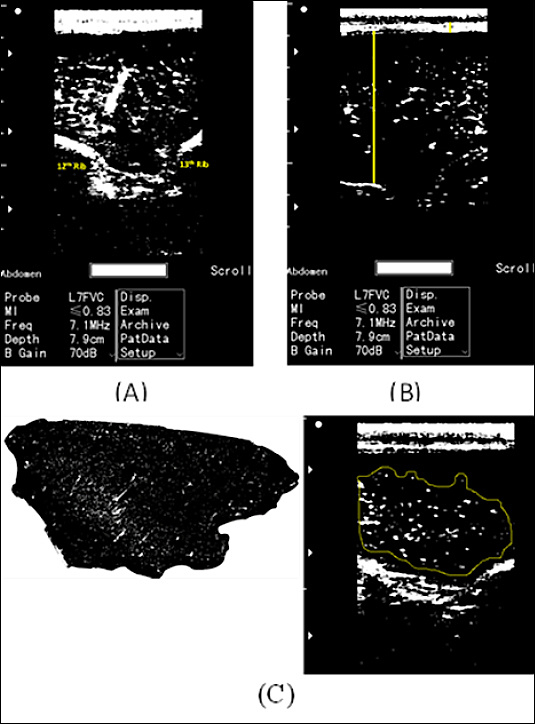
Figure 1. Ultrasound image analysis using ImageJ software in live Bali cattle. (A) Ribeye ultrasound image obtained between the 12th and 13th ribs. (B) Longitudinal ultrasound image used to measure backfat thickness and
FA methyl ester preparation
Approximately 250 g of tenderloin muscle was collected from the right side of each carcass about 1 h post-mortem and stored at −20°C. From this sample, 40 g was used for FA analysis. Lipids were extracted using a chloroform–methanol solution, followed by transesterification to produce fatty acid methyl esters (FAMEs). FAMEs were extracted, centrifuged, dried in hexane, resuspended in chloroform to remove impurities, and purified by solid-phase extraction.
Gas chromatography–flame ionization detector conditions
FAMEs (1 μL) were analyzed using gas chromatography equipped with a flame ionization detector. Separation was achieved on a cyanopropyl methyl sil capillary column (60 m × 0.25 mm, 0.25 μm film thickness) with nitrogen as the carrier gas at 30 mL/min. Injector and detector temperatures were set at 220°C and 240°C, respectively. The oven temperature program was 125°C for 5 min, then increased to 225°C at 10, 5, and 3°C/min, with holding times of 5, 10, and 7 min, respectively. The split ratio was 1:80, the injection volume was 1 μL, and the linear velocity was 23.6 cm/s. FAMEs were quantified as relative percentages using a mixed FAME standard for retention-time and peak-area comparisons. Quality control was ensured by standard injections before sample analysis. Fatty acids were grouped into saturated fatty acids (SFA), monounsaturated fatty acids (MUFA), and polyunsaturated fatty acids (PUFA) [27]. Total unsaturated fatty acids (UFA) were calculated as the sum of MUFA and PUFA. Analyses were performed in duplicate, and instruments were calibrated according to the manufacturer’s guidelines.
Population genetics and SNP diversity analysis
Allele and genotype frequencies, observed and expected heterozygosity, and Hardy–Weinberg equilibrium were calculated using PopGen 1.32 software. Polymorphic information content (PIC) was calculated using the formula:
where pi and pj represent allele frequencies at a given locus.
Statistical analysis
Associations between
Yij = μ + Gi + eij,
where Yij is the phenotypic observation, μ is the overall mean, Gi is the genotype effect, and eij is the random error. Carcass and meat traits, including FA composition, were corrected to 36 months of age, body weight, and similar environmental conditions using the equation:
Xi(corrected) = (X̅standard / X̅observation) × Xi(observation)
where Xi(corrected) is the corrected value, X̅standard is the standard group mean, X̅observation is the observation group mean, and Xi(observation) is the observed value.
RESULTS
FABP4 polymorphism
Genetic variation in

Figure 2. Partial sequencing maps of the
Table 1. SNP information of the
| Gene | SNP | Location | Variation type | dbSNP | Amino acids |
|---|---|---|---|---|---|
|
| g.4631T>C | Intron 3 | Transition | Novel | – |
|
| g.4724T>C | Intron 3 | Transition | Novel | – |
|
| g.4769G>A | Intron 3 | Transition | Novel | – |
|
| g.5002C>T | Exon 4 | Transition | Novel | Val/Ala |
Ala = Alanine, SNP = Single-nucleotide polymorphism, Val = Valine.
Table 2. Allelic and genotypic frequencies and diversity parameters of
| Gene | SNP | N | AA | AB | BB | A | B | Ho | He | χ² test | PIC |
|---|---|---|---|---|---|---|---|---|---|---|---|
|
| g.4631T>C | 95 | 0.00 | 0.08 | 0.92 | 0.04 | 0.96 | 0.084 | 0.081 | 0.160 ns | 0.077 |
|
| g.4724T>C | 95 | 0.93 | 0.07 | 0.00 | 0.96 | 0.04 | 0.074 | 0.071 | 0.119 ns | 0.068 |
|
| g.4769G>A | 95 | 0.78 | 0.21 | 0.01 | 0.88 | 0.12 | 0.211 | 0.206 | 0.051 ns | 0.184 |
|
| g.5002C>T | 95 | 0.84 | 0.14 | 0.02 | 0.91 | 0.09 | 0.137 | 0.164 | 2.721 ns | 0.150 |
AA = Reference genotype (wild-type), AB = Heterozygous genotype, BB = Mutant genotype,
Genetic association of FABP4 variants with meat quality traits
The associations between
Table 3. Association of
| SNP | Genotype (N) | LDT (mm) | BFT (mm) | MS | IMF (%) |
|---|---|---|---|---|---|
| g.4631T>C | TC (8) | 47.24 ± 6.11 | 1.84 ± 0.30 | 1.36 ± 0.19 | 2.22 ± 0.47 |
| CC (83) | 47.16 ± 5.77 | 1.88 ± 0.31 | 1.51 ± 0.56 | 2.58 ± 1.39 | |
| g.4724T>C | TT (84) | 47.23 ± 5.76 | 1.89 ± 0.31 | 1.51 ± 0.56 | 2.57 ± 1.39 |
| TC (7) | 46.51 ± 6.21 | 1.79 ± 0.30 | 1.39 ± 0.19 | 2.28 ± 0.48 | |
| g.4769G>A | GG (70) | 46.71 ± 5.12 | 1.89 ± 0.32 | 1.51 ± 0.57 | 2.57 ± 1.41 |
| GA (20) | 49.36 ± 6.99 | 1.84 ± 0.28 | 1.42 ± 0.36 | 2.35 ± 0.91 | |
| AA (1) | 35.17 ± nc | 1.54 ± nc | 2.61 ± nc | 5.30 ± nc | |
| g.5002C>T | CC (77) | 46.61b ± 5.73 | 1.86 ± 0.31 | 1.51 ± 0.57 | 2.57 ± 1.43 |
| CT (12) | 51.18a ± 4.42 | 2.00 ± 0.23 | 1.39 ± 0.23 | 2.28 ± 0.56 | |
| TT (2) | 44.67ab ± 7.40 | 1.77 ± 0.57 | 1.76 ± 0.41 | 3.20 ± 1.03 |
BFT = Backfat thickness,
Genetic association of FABP4 variants with fatty acid composition
The relationship between
Table 4. Association of
| Fatty acid (%) | g.4631T>C TC (1) | g.4631T>C CC (43) | g.4724T>C TT (43) | g.4724T>C TC (1) | g.4769G>A GG (35) | g.4769G>A GA (9) | g.5002C>T CC (40) | g.5002C>T CT (4) |
|---|---|---|---|---|---|---|---|---|
| Fat content | 3.57 ± nc | 3.18 ± 1.23 | 3.18 ± 1.23 | 3.57 ± nc | 3.17 ± 1.22 | 3.26 ± 1.27 | 3.26 ± 1.22 | 2.43 ± 0.92 |
| C8:0 | 0.00 ± nc | 0.06 ± 0.20 | 0.06 ± 0.20 | 0.00 ± nc | 0.06 ± 0.21 | 0.04 ± 0.11 | 0.06 ± 0.20 | 0.07 ± 0.14 |
| C12:0 | 0.18 ± nc | 0.07 ± 0.02 | 0.07 ± 0.02 | 0.18 ± nc | 0.07 ± 0.03 | 0.07 ± 0.01 | 0.07 ± 0.03 | 0.06 ± 0.02 |
| C13:0 | 0.04 ± nc | 0.03 ± 0.02 | 0.03 ± 0.02 | 0.04 ± nc | 0.03 ± 0.02 | 0.03 ± 0.01 | 0.03 ± 0.02 | 0.03 ± 0.03 |
| C14:0 | 3.35 ± nc | 2.17 ± 0.53 | 2.17 ± 0.53 | 3.35 ± nc | 2.15 ± 0.55 | 2.37 ± 0.56 | 2.21 ± 0.58 | 2.00 ± 0.27 |
| C14:1 | 0.09 ± nc | 0.28 ± 0.37 | 0.28 ± 0.37 | 0.09 ± nc | 0.28 ± 0.38 | 0.26 ± 0.31 | 0.29 ± 0.38 | 0.18 ± 0.29 |
| C15:0 | 0.81 ± nc | 0.59 ± 0.36 | 0.59 ± 0.36 | 0.81 ± nc | 0.61 ± 0.37 | 0.55 ± 0.26 | 0.60 ± 0.37 | 0.52 ± 0.07 |
| C16:0 | 32.95 ± nc | 20.82 ± 3.61 | 20.82 ± 3.61 | 32.95 ± nc | 21.10 ± 4.09 | 21.05 ± 3.89 | 21.16 ± 4.17 | 20.43 ± 1.69 |
| C16:1 | 2.00 ± nc | 1.30 ± 0.35 | 1.30 ± 0.35 | 2.00 ± nc | 1.31 ± 0.38 | 1.33 ± 0.31 | 1.33 ± 0.37 | 1.19 ± 0.12 |
| C17:0 | 2.13 ± nc | 1.99 ± 0.77 | 1.99 ± 0.77 | 2.13 ± nc | 2.01 ± 0.83 | 1.92 ± 0.42 | 2.00 ± 0.76 | 1.96 ± 0.84 |
| C17:1 | 0.44 ± nc | 0.26 ± 0.14 | 0.26 ± 0.14 | 0.44 ± nc | 0.27 ± 0.14 | 0.26 ± 0.16 | 0.27 ± 0.14 | 0.22 ± 0.17 |
| C18:0 | 36.55 ± nc | 31.91 ± 3.64 | 31.91 ± 3.64 | 36.55 ± nc | 32.00 ± 4.01 | 32.09 ± 2.01 | 32.11 ± 3.65 | 31.05 ± 4.32 |
| C18:1n9c | 0.10 ± nc | 12.45 ± 4.51 | 12.45 ± 4.51 | 0.10 ± nc | 11.64 ± 5.00 | 14.21 ± 3.65 | 12.06 ± 5.04 | 13.25 ± 1.70 |
| C18:1n9t | 1.72 ± nc | 3.03 ± 1.25 | 3.03 ± 1.25 | 1.72 ± nc | 2.86 ± 1.25 | 3.56 ± 1.17 | 3.07 ± 1.28 | 2.27 ± 0.71 |
| C18:2n6c | 3.88 ± nc | 1.75 ± 0.76 | 1.75 ± 0.76 | 3.88 ± nc | 1.83 ± 0.91 | 1.67 ± 0.23 | 1.75 ± 0.78 | 2.27 ± 1.18 |
| C18:3n3 | 0.00 ± nc | 0.23 ± 0.27 | 0.23 ± 0.27 | 0.00 ± nc | 0.21 ± 0.27 | 0.28 ± 0.26 | 0.21 ± 0.26 | 0.41 ± 0.29 |
| SFA | 77.19 ± nc | 58.56 ± 5.99 | 58.56 ± 5.99 | 77.19 ± nc | 58.98 ± 7.05 | 59.01 ± 4.42 | 59.16 ± 6.60 | 57.16 ± 6.66 |
| MUFA | 10.40 ± nc | 20.31 ± 4.32 | 20.31 ± 4.32 | 10.40 ± nc | 19.47 ± 4.50 | 22.48 ± 4.02 | 20.04 ± 4.68 | 20.61 ± 2.90 |
| PUFA | 5.14 ± nc | 17.65 ± 4.75 | 17.65 ± 4.75 | 5.14 ± nc | 16.71 ± 5.12 | 19.91 ± 4.11 | 17.38 ± 5.27 | 17.27 ± 2.30 |
| UFA | 5.26 ± nc | 2.66 ± 1.09 | 2.66 ± 1.09 | 5.26 ± nc | 2.76 ± 1.26 | 2.57 ± 0.51 | 2.66 ± 1.12 | 3.34 ± 1.34 |
| Total FA | 87.60 ± nc | 78.66 ± 7.03 | 78.66 ± 7.03 | 87.60 ± nc | 78.33 ± 7.20 | 80.96 ± 6.51 | 79.01 ± 7.10 | 77.44 ± 7.67 |
DISCUSSION
Genetic diversity and population structure
Genetic diversity is a key determinant of adaptability and long-term sustainability in livestock populations. Based on allele and genotype frequency data, the AA genotype was more frequently observed for SNPs g.4724T>C, g.4769G>A, and g.5002C>T, whereas the BB genotype predominated for g.4631T>C (Table 2). Adequate genetic variation is essential for breeding programs aimed at improving performance and ensuring food security [28, 29]. Preservation of genetic diversity reduces the risks associated with inbreeding and genetic uniformity, thereby maintaining resilience, fitness, and protection against inbreeding depression [30].
All SNPs identified in this study were polymorphic, with major allele frequencies <99% and minor allele frequencies >1% [31]. The observed heterozygosity (Ho) was 50% for all SNPs, indicating relatively low heterozygosity in the Bali cattle population. This pattern may reflect inbreeding, genetic drift, and a small effective population size, which can increase homozygosity and allele fixation [32]. Reduced genetic variation may negatively affect overall health and adaptability, increasing susceptibility to diseases and environmental challenges [30]. All SNPs conformed to Hardy–Weinberg equilibrium, suggesting stable allele and genotype frequencies that were not significantly influenced by selection, migration, mutation, or genetic drift [33]. This equilibrium indicates population stability and limited disturbance from breeding practices or environmental pressures [34]. In addition, all SNPs in
Association of FABP4 polymorphisms with meat quality traits
The
Previous studies have reported significant associations between
Association of FABP4 polymorphisms with fatty acid composition
The absence of significant associations between
Other studies have reported significant relationships between
Implications for marker-assisted selection
The SNP g.5002C>T in
CONCLUSION
This study identified four novel SNPs in the
The significant association between
A major strength of this work lies in the integration of molecular genetics with
Despite these strengths, the study was conducted on a single population with a relatively limited sample size, which may have constrained the detection of associations for traits with low genetic variance or unbalanced genotype frequencies. The low heterozygosity observed for several SNPs may also have reduced statistical power, particularly for traits such as MS, IMF, and FA composition that are strongly influenced by multiple genes and environmental factors.
Future studies should validate the effect of
In conclusion, this study provides the first evidence that
DATA AVAILABILITY
All the generated data are included in the manuscript.
AUTHORS’ CONTRIBUTIONS
DD and JJ: Conceptualization of the study, investigation, methodology, data analysis, laboratory work, interpretation of data, and drafted, reviewed, and edited the manuscript. DD, MFU, and JJ: Collected the animal sample and performed ultrasound imaging. SS, AF, and IK: Validation, investigation, and reviewed and edit the manuscript. CS: Data analysis and revised the manuscript. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
This research was funded by PMDSU program (contract number: 3677/IT3/L1/PT/01.03/P/B/2022) years 2022 from the Ministry of Research and Technology or granted by the Ministry of Education and Culture Republic of Indonesia (by Dairoh); Postdoctoral Fellowship at the Research Center for Applied Zoology, National Research and Innovation Agency (BRIN), Indonesia No: B-3464/III.5/SI.06/6/2025 (By Dairoh), and Penelitian Fundamental Reguler (PFR) from the Ministry of Education, Culture, Research, and Technology No: 006/C3/DT.05.00/PL/2025 (by Jakaria). The authors would like to express their gratitude to the field personnel for their support and to the National Research and Innovation Agency’s Talent Management for providing the postdoctoral program.
REFERENCES
- Desmukh SS, Elayadeth-Meethal M, Abraham BL, Asaf M, Lali FA. Sustainable livestock production:Prospects for innovation. Hyderabad: MANAGE; 2023. p. 111. [Google Scholar]
- Vázquez-Mosquera JM, Fernandez-Novo A, de Mercado E, Vázquez-Gómez M, Gardon JC, Pesántez-Pacheco JL. Beef nutritional characteristics, fat profile and blood metabolic markers from purebred Wagyu, crossbred Wagyu and crossbred European steers raised on a fattening farm in Spain. Animals 2023;13(5):846. [Google Scholar] | [Crossref]
- Baik M, Lee J, Kim SY, Ranaweera KKTN. Factors affecting beef quality and nutrigenomics of intramuscular adipose tissue deposition. Anim Biosci 2023;36(2):350-363. [Google Scholar] | [Crossref]
- Inoue K, Shoji N, Hinda T, Oyama K. Genetic relationships between meat quality traits and fatty acid composition in Japanese Black cattle. Anim Sci J 2017;88(1):11-18. [Google Scholar] | [Crossref]
- Martojo H. Indigenous Bali cattle is most suitable for sustainable small farming in Indonesia. Reprod Domest Anim 2012;47(1):10-14. [Google Scholar] | [Crossref]
- Tahuk PK, Oktavianus R, Nahak R, Gerson FB. The effect of complete feed to carcass characteristics and meat quality of male Bali cattle fattened in West Timor, Indonesia. Vet World 2020;13(11):2515-2527. [Google Scholar] | [Crossref]
- Dairoh, Ulum MF, Jakaria, Sumantri C. Calpain 1 gene expression in liver tissue and the association of novel calpain 1 single-nucleotide polymorphisms with meat quality in Bali cattle. Arch Anim Breed 2024;67((3)):311-321. [Google Scholar] | [Crossref]
- Dairoh, Sutikno, Ishak AB, Priyanto R, Sumantri C, Ulum MF, Jakaria. Fatty acid profiling of Bali and Wagyu cattle using principal component analysis. Bull Anim Sci 2024;48(1):64-69. [Google Scholar] | [Crossref]
- Sharma P, Doultani S, Hadiya KK, George LB, Highland HN. Overview of marker-assisted selection in animal breeding. J Adv Biol Biotechnol 2024;27(5):303-318. [Google Scholar] | [Crossref]
- Meuwissen T, Hayes B, Goddard M. Genomic selection:A paradigm shift in animal breeding. Anim Front 2016;6(1):6-14. [Google Scholar] | [Crossref]
- Ribeca C, Bonfatti V, Cecchinato A, Albera A, Gallo L, Carnier P. Effect of polymorphisms in candidate genes on carcass and meat quality traits in double muscled Piemontese cattle. Meat Sci 2014;96(3):1376-1383. [Google Scholar] | [Crossref]
- Josephrajan A, Hertzel AV, Bohm EK, McBurney MW, Imai SI, Mashek DG. Unconventional secretion of adipocyte fatty acid binding protein 4 is mediated by autophagic proteins in a sirtuin-1-dependent manner. Diabetes 2019;68((9)):1767-1777. [Google Scholar] | [Crossref]
- Zhang Y, Zhang J, Ren Y, Lu R, Yang L, Nie G. Tracing the evolution of fatty acid-binding proteins in organisms with heterogeneous fat distribution. FEBS Open Bio 2020;10(5):861-872. [Google Scholar] | [Crossref]
- Furuhashi M. Fatty acid-binding protein 4 in cardiovascular and metabolic diseases. J Atheroscler Thromb 2019;26(3):216-232. [Google Scholar] | [Crossref]
- Xiang J, Li H, Guo Z, Li T, Yamada T, Li X. Effect of
FABP4 gene polymorphisms on fatty acid composition, chemical composition, and carcass traits in Sonid sheep. Animals 2025;15(2):226. [Google Scholar] | [Crossref] - Blecha IM, Siqueira F, Ferreira AB, Feijó GL, Torres Junior RA, Medeiros SR. Identification and evaluation of polymorphisms in
FABP3 andFABP4 in beef cattle. GMR 2015;14(4):16353-16363. [Google Scholar] | [Crossref] - Dawood M, Kramer LM, Shabbir MI, Reecy JM. Genome-wide association study for fatty acid composition in American Angus cattle. Animals 2021;11(18):2424. [Google Scholar] | [Crossref]
- Goszczynski DE, Papaleo-Mazzucco J, Ripoli MV, Villarreal EL, Rogberg-Muñoz A, Mezzadra CA. Genetic variation in
FABP4 and evaluation of its effects on beef cattle fat content. Anim Biotechnol 2017;28(3):211-219. [Google Scholar] | [Crossref] - Nishimura T. The role of intramuscular connective tissue in meat texture. Anim Sci J 2010;81(1):21-27. [Google Scholar] | [Crossref]
- Mallick R, Basak S, Das RK, Banerjee A, Paul S, Pathak S, Duttaroy AK. Fatty acids and their proteins in adipose tissue inflammation. Cell Biochem Biophys 2024;82:35-51. [Google Scholar] | [Crossref]
- Hocquette JF, Gondret F, Baéza E, Médale F, Jurie C, Pethick DW. Intramuscular fat content in meat-producing animals:Development, genetic and nutritional control, and identification of putative markers. Animal 2010;4(2):303-319. [Google Scholar] | [Crossref]
- Listrat A, Lebret B, Louveau I, Astruc T, Bonnet M, Lefaucheur L. How muscle structure and composition influence meat and flesh quality. Sci World J 2016;2016:1-14. [Google Scholar] | [Crossref]
- Chen J, Chen F, Lin X, Wang Y, He J, Zhao Y. Effect of excessive or restrictive energy on growth performance, meat quality, and intramuscular fat deposition in finishing Ningxiang pigs. Animals 2021;11(1):27. [Google Scholar] | [Crossref]
- Bayraktar M, Goncu S, Ergul A, Karaman R, Ozcan BD, Ergul S. Association between
FABP3 andFABP4 genes with changes in milk composition and fatty acid profiles in native Southern Yellow cattle. Vet Sci 2025;12(19):893. [Google Scholar] | [Crossref] - Guidelines for uniform beef improvement program. North Carolina: North Carolina State University; 2016. p. 35-46. [Google Scholar]
- Jakaria, Khasanah H, Priyanto R, Baihaqi M, Ulum MF. Prediction of meat quality in Bali cattle using ultrasound imaging. Indones Trop Anim Agric 2017;42(2):59-65. [Google Scholar] | [Crossref]
- Dairoh, Ulum MF, Jakaria, Ishak ABL, Sumantri C. A novel SNPs of the
SREBF1 andSCARB1 genes and the association with fatty acid profile in Bali cattle. Trop Anim Sci J 2023;46(4):428-438. [Google Scholar] | [Crossref] - Blackburn HD, Wilson CS, Krehbiel B. Conservation and utilization of livestock genetic diversity in the United States of America through gene banking. Diversity 2019;11(12):244. [Google Scholar] | [Crossref]
- Martyniuk E. Policy effects on the sustainability of animal breeding. Sustainability 2021;13(14):7787. [Google Scholar] | [Crossref]
- Hoban S, Arche FI, Bertol LD, Bragg JG, Breed MF, Bruford MW. Global genetic diversity status and trends:Towards essential biodiversity variables for genetic composition. Biol Rev Camb Philos Soc 2022;97(4):1511-1538. [Google Scholar] | [Crossref]
- Allendorf FW, Luikart G, Aitken SN. Conservation and the genetics of populations. Hoboken: Wiley-Blackwell; 2013. p. 602. [Google Scholar]
- Kardos M, Taylor HR, Ellegren H, Luikart G, Allendorf FW. Genomics advances the study of inbreeding depression in the wild. Evol Appl 2016;9(10):1205-1218. [Google Scholar] | [Crossref]
- Abramovs N, Brass A, Tassabehji M. Hardy-Weinberg equilibrium in the large-scale genomic sequencing era. Front Genet 2020;11:210. [Google Scholar] | [Crossref]
- Boes J, Boettcher P, Honkatukia M. Innovations in cryoconservation of animal genetic resources. Rome: FAO; 2023. p. 41-47. [Google Scholar]
- Harish A, Lopes Pinto FA, Eriksson S, Johansson AM. Genetic diversity and recent ancestry based on whole-genome sequencing of endangered Swedish cattle breeds. BMC Genomics 2024;25(1):89. [Google Scholar] | [Crossref]
- Strucken EM, Gebrehiwot NZ, Swaminathan M, Joshi S, Al Kalaldeh M, Gibson JP. Genetic diversity and effective population sizes of thirteen Indian cattle breeds. Genet Sel Evol 2021;53(1):47. [Google Scholar] | [Crossref]
- Ardicli S, Samli H, Alpay F, Dincel D, Soyudal B, Balci F. Association of SNP in
FABP4 gene with carcass characteristics and meat quality in Holstein bulls. Ann Anim Sci 2017;17(1):117-130. [Google Scholar] | [Crossref] - Gao YY, Cheng G, Cheng ZX, Bao C, Yamada T, Cao GF. Association of variants in
FABP4 , FASN, SCD, SREBP1 and TCAP genes with intramuscular fat and carcass traits in Qinchuan cattle. Meat Sci 2022;192:108882. [Google Scholar] | [Crossref] - Mwangi FW, Pewan SB, Otto JR, Adegboy O, Charmley E, Gardiner CP. SNPs in
FABP4 , FASN and SCD genes influence carcass traits of tropical crossbred beef steers. Agriculture 2022;12(8):1171. [Google Scholar] | [Crossref] - Shin SC, Heo JP, Chung ER. Genetic variants of
FABP4 gene associated with marbling score and meat quality in Hanwoo cattle. Mol Biol Rep 2012;39(5):5323-5330. [Google Scholar] | [Crossref] - Serdar CC, Cihan M, Yücel D, Serdar MA. Sample size, power and effect size revisited. Biochem Med 2021;31((1)):010502. [Google Scholar] | [Crossref]
- Hoashi S, Hinenoya T, Tanaka A, Ohsaki H, Sasazaki S, Taniguchi M. Association between fatty acid composition and
FABP4 and LXR-alpha genotypes in Japanese Black cattle. BMC Genet 2008;9(1):84. [Google Scholar] | [Crossref] - Bergman BC, Howard D, Schauer IE, Maahs DM, Snell-Bergeon JK, Clement TW. Importance of palmitoleic acid to insulin sensitivity. J Clin Endocrinol Metab 2013;98(1):E40-E50. [Google Scholar] | [Crossref]
- Bermúdez MA, Pereira L, Fraile C, Valerio L, Balboa MA, Balsinde J. Roles of palmitoleic acid and positional isomers in inflammation and metabolic diseases. Cells 2022;11(14):2146. [Google Scholar] | [Crossref]
- Anzhany D, Toharmat T, Despal, Lozicki A. Fatty acid biohydrogenation, fermentation, and digestibility of ration containing napier and king grass with different harvest ages and altitudes:
in vitro study. Trop Anim Sci J 2023;47(1):68-78. [Google Scholar] | [Crossref] - Mercola J, D'Adamo CR. Linoleic acid and chronic disease associations. Nutrients 2023;15(14):3129. [Google Scholar] | [Crossref]
- Yin BZ, Fang JC, Zhang JS, Zhang LM, Xu C, Xu H. SNPs in
FABP4 correlate with meat quality and lipid metabolism in Yanbian yellow cattle. PLOS One 2020;15(6):1-14. [Google Scholar] | [Crossref] - Oh D, Lee Y, La B, Yeo J. SNP identification for fatty acid composition associated with FABP4 in Korean cattle. Asian-Australas J Anim Sci 2012;25(7):913-920. [Google Scholar] | [Crossref]
- Kostusiak P, Slósarz J, Puppel K, Gołębiewski M, Grodkowski G. Polymorphism of genes and impact on beef quality. Curr Issues Mol Biol 2023;46(6):4749-4762. [Google Scholar] | [Crossref]
- Sakowski T, Grodkowski G, Gołębiewski M, Puppel K, Kostusiak P, Slósarz J. Genetic and environmental determinants of beef quality. Front Vet Sci 2022;9:819605. [Google Scholar] | [Crossref]
- Neumann GB, Korkuć P, Arends D, Wolf MJ, May K, Reißmann M. Design and performance of bovine 200k SNP chip for endangered cattle. BMC Genomics 2021;22(1):905. [Google Scholar] | [Crossref]